��
��
Natural killer cells and autoimmunity
��
Natural Killer Cells
PDF icon Download Natural Killer Cells.pdf (419.94 KB)
Philipp Eissmann, Imperial College, London, UK
Natural Killer (NK) Cells are lymphocytes in the same family as T and B cells, coming from a common progenitor. However, as cells of the innate immune system, NK cells are classified as group I Innate Lymphocytes (ILCs) and respond quickly to a wide variety of pathological challenges. NK cells are best known for killing virally infected cells, and detecting and controlling early signs of cancer. As well as protecting against disease, specialized NK cells are also found in the placenta and may play an important role in pregnancy.
NK cells were first noticed for their ability to kill tumour cells without any priming or prior activation (in contrast to cytotoxic T cells, which need priming by antigen presenting cells). They are named for this ��natural�� killing. Additionally, NK cells secrete cytokines such as IFN�� and TNF��, which act on other immune cells like Macrophage and Dendritic cells to enhance the immune response.
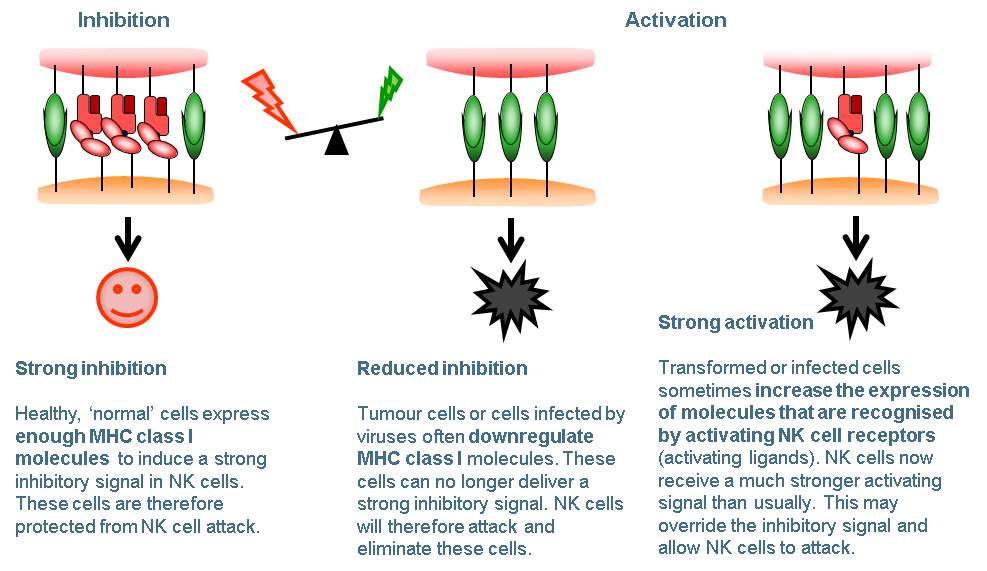
While on patrol NK cells constantly contact other cells. Whether or not the NK cell kills these cells depends on a balance of signals from activating receptors and inhibitory receptors on the NK cell surface. Activating receptors recognise molecules that are expressed on the surface of cancer cells and infected cells, and ��switch on�� the NK cell. Inhibitory receptors act as a check on NK cell killing. Most normal healthy cells express MHC I receptors which mark these cells as ��self��. Inhibitory receptors on the surface of the NK cell recognise cognate MHC I, and this ��switches off�� the NK cell, preventing it from killing. Cancer cells and infected cells often lose their MHC I, leaving them vulnerable to NK cell killing. Once the decision is made to kill, the NK cell releases cytotoxic granules containing perforin and granzymes, which leads to lysis of the target cell.
Natural killer cells - figure 1
The genes for both MHC I and the NK cell inhibitory receptors which recognise them vary a lot between individuals. The versions (or alleles) of these genes a person carries have been linked to their ability to fight HIV infection and their risk of some autoimmune diseases. NK cell varieties also change with age and are affected by chronic viral infections such as cytomegalovirus (CMV).
Because of their ability to kill tumour cells, NK cells are an attractive target for cancer immunotherapies. Some therapeutic monoclonal antibodies rely on NK cell killing. Researchers are also developing treatments to activate NK cells using small molecules or cytokines, and even testing genetically modified living NK cells as therapies
Natural Killer Cells | British Society for Immunology
https://www.immunology.org/public-information/bitesized-immunology/cells/natural-killer-cells��
��֢����������������������̳ҽԺ�о��ƣ������50�����ܰ�ϸ�����Խ��͵��¹ڷ��ף�Ӧ����������֢���
�й�ѭ����־
02-13 23:17
2��12�գ���ҽ�ƴ�ѧ����������̳ҽԺ�о�������ϸ��/�ܰ�ϸ����ֵ��Ա��ҽѧ����������Ԥӡ��ƽ̨medRvix������һ���о���ʾ��������ϸ��/�ܰ�ϸ����ֵ��NLR�����������ڷ�����֢����״�������ף��¹ڷ��ף����ߡ�
�о��߷��֣������50����NLR��3.13�Ļ��߽�չΪ��֢�Ŀ����Դ�Ӧ����ת����֢��������ơ�
�������50��Ļ����У�NLR��3.13��һ����չΪ��֢����NLR<3.13�Ļ����н�9.1%��չΪ��֢��
�о�����Ϊ�������¹ڷ����ߣ����Ը���NLR���������Σ�շֲ������
����<50����NLR<3.13�Ļ��߽�չΪ��֢�ķ���Ϊ0������������ҽԺ����и��롣
����<50�ꡢNLR��3.13�Ļ��߽�չΪ��֢�ķ��սϵͣ���Ҫס��ͨ���벡�����ơ�
�����50�ꡢNLR<3.13�Ļ��߽�չΪ��֢�ķ������еȵģ�Ӧ��Ժ�������ƣ������к�������֧�����ơ�
�����50�ꡢNLR��3.13�Ļ��߽�չΪ��֢�ķ��սϸߣ�Ӧ�û���ת����֢������������д�����ϵͳ֧�֡�
�о�����Ϊ�����д�������������£�����Σ�շֲ�������������ڻ���ҽ����Դ��ȱ��������֢���ߵ������ʡ�
���о�ǰհ�����뱱����̳ҽԺ��2020��1��13����1��31�����ε�61���¹ڷ����ߣ�����LASSO COX�ع������֢���ߵ�Ԥ�����ء�
���������ʾ��NLR���¹ڷ����߽�չΪ��֢�Ķ���Σ�����أ�����Ԥ��ȷ�Խϸߣ�cָ��Ϊ0.807����
��Դ��Neutrophil-to-Lymphocyte Ratio Predicts Severe Illness Patients with 2019 Novel Coronavirus in the Early Stage. medRvix, Posted February 12, 2020.��
Neutrophil in Viral Infections, Friend or Foe?
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3557572
Polymorphonuclear leukocytes or neutrophils are the first immune cells to the site of injury and microbial infection. Neutrophils are crucial players in controlling bacterial and fungal infections, and in particular secondary infections, by phagocytosis, degranulation and neutrophil extracellular traps (NETs).
Cited by: 61
Publish Year: 2013
Author: Brandon D. Drescher, Fengwei Bai
A Role for Neutrophils in Viral Respiratory Disease
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5427094
May 12, 2017 �� Thus, there is evidence that the role of neutrophils in viral infections of the respiratory system is not limited to inflammation, but likely includes recovery from infection and the initiation of adaptive immunity (74, 138, 284).
Cited by: 29
Publish Year: 2017
Author: Jeremy V. Camp, Colleen B. Jonsson��
CIK��cytokine-induced killer������������[����ϸ�������Ʒ�]����ϸ�������յ���ɱ��ϸ��
CIK��cytokine-induced killer������������[����ϸ�������Ʒ�]����ϸ�������յ���ɱ��ϸ�������ڸ���ϸ��ͬʱ����CD3+��CD56+����Ĥ�����ӣ����ֱ���ΪNKϸ����T�ܰ�ϸ���������T�ܰ�ϸ��ǿ��Ŀ������Ժ�NKϸ���ķ�MHC������ɱ���ŵ㡣��ˣ�Ӧ��CIKϸ������Ϊ����һ������������ϸ���������Ƶ���ѡ������CIKϸ���е�ЧӦϸ��CD3+��CD56+ϸ��������������Ѫ�н��٣���1%-5%��
����
��νCIKϸ�������ǽ��˵�����Ѫ������ϸ���������ö���ϸ������[�翹CD3����(CD3McAb)����ϸ������-2(IL-2)��������-�á� (IFN -��)��IL��1����]��ͬ����һ��ʱ����õģ����з���Ҫ��֯�����Կ�ԭ(MHC)������ɱ�����Ե�һȺ����ϸ���� [1]
�ص�
CIKϸ���е�ЧӦϸ��CD3+CD56+ϸ��������������Ѫ�м��亱������1%��5%�������⾭����������28��30�죬CD3+CD56+ϸ��Ѹ�����࣬������ǰ�����ɴ�1000�����ϡ� [2] ʵ��֤������������CD3+CD56+ϸ����Դ��CD3+CD56-Tϸ������CD3-CD56+NKϸ����ͬʱ������CD3+CD56-��T ϸ���У���CD4+CD8-Tϸ���⣬��������T ϸ����Ⱥ��CD4-CD8+��CD4-CD8-��CD4+CD8+������ͨ��������������������CD56���ӵı��������CD4+CD8+ϸ����CD4-CD8-ϸ��������������Ѫ�к������Ͷ������ʾ��CD3+CD56+ϸ�����������Դ������Ѫ��CD4-CD8+Tϸ����������CD4-CD8-Tϸ��������1���º��н�56%��T ϸ��ͬʱ����CD56��CD3��������Ҳ��CIKϸ������Ҫ��Դ���Ƚ�CD3+CD56+CIKϸ���б���CD8+��CD8-������Ⱥϸ����ɱ������û�������Բ��죬��ʾCIKϸ����ϸ��������CD3CD56�����������ƣ�����CD8�ı���δ���ֳ�����ԡ�
1.CIKϸ����ֳ�ٶȿ죬����������ϸ���ɴ�����ֳ����ϸ������Ҳ�����ǿ��
2.CIKϸ������ʶ�������Ļ��ƣ���������ϸ���������á�
3.ɱ���㣬�����ڰ�Ѫ�����ܰ������ΰ���θ���������ȶ������������ƣ��Զ�����ҩ����ϸ��ͬ�����С�
4.�ǵ��͵ĸ��Ի���������ģʽ��������ϸ���������ʹ��������������ߣ���������Ŀ��������ã��Ӷ�����������ʩ��˫�ص����á�
5.����CIKϸ���ǻ������ϸ�����������dz���ȫ��
ɱ��ԭ��
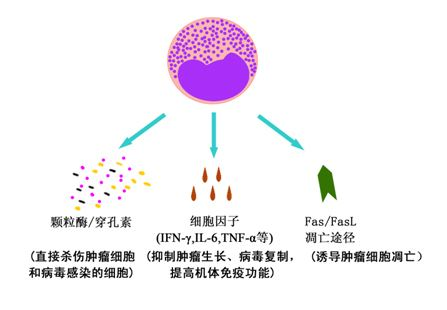
CIKϸ���ܹ�ͨ������;��ɱ������ϸ���Ͳ�����Ⱦϸ����
��CIKϸ��������ϸ���Ͳ�����Ⱦϸ����ֱ��ɱ�ˣ�CIKϸ������ͨ����ͬ�Ļ���ʶ������ϸ�����ͷſ���ø/�����صȶ��Կ�������������ϸ���ѽ⡣
��CIKϸ���ͷŵĴ�������ϸ�����Ӿ�������ɱ�����ԣ�����������CIKϸ�����Է��ڶ���ϸ�����ӣ���IFN-�á�TNF-����IL-2�ȣ�����������ϸ����ֱ���������ã�����ͨ�����ڻ�������ϵͳ��Ӧ�Լ��ɱ������ϸ����
��CIKϸ���ܹ��յ�����ϸ���ĵ�����CIKϸ�������������б���FasL�����Ϳ�Ĥ�ǵ��ף�ͨ��������ϸ��Ĥ�����Fas�����Ϳ�Ĥ�ǵ��ף���ϣ��յ�����ϸ��������
CIKϸ���������õ�����;��
CIKϸ���������õ�����;��
ɱ���ص�
Ӧ��LAKϸ���ǵ����Ϊ�ռ������������������Ʒ������㷺ʹ���ں�ɫ��������ϸ�������ǻ�����ܰ������ΰ��ͽ᳦������LAKϸ�������������ޣ�ɱ������Ҳ��TIL��T�ܰ�ϸ��Ϊ�ͣ�����Ȼɱ���㣬��Ч�����ޡ���Ƚ϶��ԣ�TIL ϸ��������LAKϸ�����и�ǿ��Ŀ���Ч����������ɱ����խ���Ʊ����ѣ������ռ������п��ܵ��µĹ��ܸı�����������ٴ�Ӧ�ü�ֵ������������ЧӦϸ����������������ȣ�CIKϸ�����ж��ص����ƣ��о����¡�
1. ��ֳ�ٶȿ�
CIKϸ�������������м���IFN-�á�IL-1������CD3��McAb��IL-2�ȶ����Ӻ�ϸ����ֳ�ٶ�Ѹ�ټӿ죬Զ����LAKϸ������������22����ֳ���ߴﶥ�壬Լ����100��������CD3+CD56+ϸ������������������1000�����ϣ�����ռ�ٷֱ�Ҳ���������������28��30��ʱ��ƽ̨�ڣ�ϸ������������ֵ����LAKϸ������ǰ������û���������ӡ�
2. ɱ�����Ը�
CIKϸ������CD3+CD56+Tϸ��Ϊ��������ϸ��Ⱥ������������ʵ��֤ʵCIKϸ������NKϸ��Ϊ����LAKϸ���߱���ǿ���ɱ�����ԣ�����������ɱ��ϸ�����Ե�ά�ֲ��������������Դ��IL-2�ij������衣����ʵ���У��λ���Lu�Ⱦ������������������CIKϸ����LAKϸ��������ϸ��ϵ��ɱ�������Ը����������CIKϸ��������������CD3+CD56+ЧӦϸ������Ѹ�٣���CIKϸ������ɱ�˵�λ��TLU��ΪLAK ϸ����73���������ߣ���ɱ��Ч����������LAKϸ����������¡����ʵ����ʾ��CIKϸ������ϸ������Logָ��Ϊ2.5��3.5����LAKϸ������ϸ������ָ����2��Log������ʵ�鷢�֣�������������������ȱ��С�����Ϲ���������B ϸ���ܰ���SU-DHL4ģ�ͣ�CIKϸ����������������������������ת�ƣ��ӳ������ڵȷ�������þ���������LAKϸ����
3. ɱ����
CIKϸ����Ȼ��CD3+CD56+ Tϸ��Ϊ��ҪЧӦϸ������ȴû��T �ܰ�ϸ��ɱ��ʱ��MHC �����ԣ��ʶ��ڶ�������ϸ��ϵ������NK���е�K562��NK�����е�Hela��HL60����T ϸ�����ܰ�Ѫ��ϸ��ϵOCRF-CEM�����ܰ���ϸ��ϵOCI-LY8��LAM53���˽᳦��ϸ��ϵHT-29��CR75��������ϸ��ϵA704��������������֯�����ֳ�ǿ���ɱ�˻��ԡ�
4. �Զ�����ҩ����ϸ��ͬ������
Wolf�ð�ù�غͳ����¼��յ���������ҩϸ��ϵK562/DOX��CCRF-CEM-VBL������CIKϸ���Ի���ҩ�����е��ױ�ϸ���Ͳ����е�ת��ϸ��������ǿ���ɱ�˻��ԣ����߱Ƚ����
5. ɱ�����Բ���CsA��FK506���������Ƽ���Ӱ��
Mehta�۲쵽�������Ƽ�CsA��FK506��Ȼ�������ƿ�CD3�����鵼��CIKϸ���ѿ������̣�ȴ��Ӱ���ϸ���յ���CIKϸ���ѿ���������CIKϸ����ϸ����ɱ�˻��Բ�����˽��͡�
6. ������������Ѫǰ��ϸ�����Ժ�С
Seheffoldͨ��CFU-GM�γ�ʵ����CIKϸ���Թ�����Ѫǰ��ϸ����Ӱ�죬����CIKϸ����K562ϸ����ɱ��ǿ�ȸߴ�3��������GM-CFU���в���1�������ơ�
HolyeҲ֤ʵCIKϸ����������ϵ��¡���ɼ���û��Ӱ�죬ֻ�ǶԺ�ϵ��������ʾ��������ƣ��������CIKϸ���������ڽϸ�ˮƽ��IFN-���йء�
7. �ֿܵ�����ϸ��������ЧӦϸ��Fas-FasL����
��֤��������������ʧ�ܵ�һ����Ҫԭ���ǹ���ЧӦϸ��������ϸ����������ijЩ���ף���Ҫ��FasL���յ���������CIKϸ����Ȼ��Fas��ռ�ݺ����������ϸ��������������ɱ��ϸ������û������Ӱ�졣Verneris��ʵ����ʾCIKϸ�����п�������������������ֱ���������cFLIP��Bcl-2��Bcl-X1��DAD1��survivin��ת¼ˮƽ�ϵ���ͬʱ����CIKϸ���߱��ϳ�FasL��������CIKϸ�����������п��Լ�������ѧ���Ե�ˮ����FasL������CIKϸ�����ԶԿ�����FasL ����������������ЧӦϸ�������½�������ʧ����
Ӱ������
1.��Դ��ϸ�����ӵIJ���
CIKϸ��������������Ҫ��Դ��ϸ�����ӣ���IL-2��IL-7��IL-12�ȵĸ�������Щ���ӿ�����������ϵͳ�ڸ��ֿ�ԭ������ϸ����������������ѧ���ԡ���Դ��IL-2��IL-7��IL-12���������ٽ��ܰ�ϸ�������������������IL-2��IL-7������CIKϸ������ֳ��Ϊ�ߣ�����Դ�ԣ�IL-2��IL-7��IL-12��CIKϸ����ϸ��������û��Ӱ�졣��Դ��IL-2��IL-7�Ĵ̼��ή��CIKϸ��������Ӧ����ı���������CD28������IL-7���������½�IL-2ʱ������ߡ�IL-12�ή��CIKϸ������ICAM-1�ı��IL-7������CD56�ı����IL-2��ȣ�IL-7����������CD4+ϸ���ı�������Ȼ��Դ��IL-2��IL-7��IL-12���������о��ɳ�����������ϸ���������о���ʾ��Դ��IL-12�����ӻ�����CIKϸ���л���ϸ���ı�����
��CD3McAb������CIKϸ������������������Ҫ���ã������CIKϸ����Ѫ�����ܰ�����ɱ����������ͬ�����дٽ����á�Lefterov����CD3McAb���ϸ��Ԥ�ȹ�ͬ����������CIKϸ�������ɱ�������ԣ�����������ǿ���ÿ��Ա���FcR�Ŀ��壨�翹CD36����CD32����������ϣ����֤����CD3McAb������ɱ�˻���������FcR�鵼�Ŀ������йء�
2.����ϸ�����ӻ����תȾ
����CIKϸ����������Դ��ϸ�����������������ͨ������ת�Ʒ�������ػ���ת��CIKϸ���������ɼ�����Դ��ϸ�����ӵ�ʹ�������������CIKϸ�������Ŀ������ԡ�
IL-7��Fitke���øĽ����ٲ���ת����ϵͳ����IL-7����תȾCIKϸ��������תȾ��ϸ���������ɽϸ�Ũ��IL-7������߿ɴ�1 100pg/106cell/24h���ϳɵ�IL-7�������Ե�����ѧ���ԣ����Դٽ�תȾCIKϸ������ֳ����������δתȾϸ������ԴIL-7����ı���ͬʱ�ı���CIKϸ��������ϸ�����ӵķ��ڣ���������TNF-�������������ߣ���һ������δתȾCIK��������IL-7ʱ��δ�۲����ȻתȾ��CIKϸ�����������ϸ��ɱ�˻�����صı��濹ԭ����ICAM-1 ����δתȾCIKϸ����������Ա仯����תȾ��CIKϸ���ڶԶ�������ϸ��ϵ�����������Ժ�ɫ�����Լ��᳦����ɱ��������δתȾCIKϸ����������ǿ��
IL-2��Lu�ȷ���CIKϸ������������CD56���ӵı�����IL-2�����Եģ�������IL-2�Ĵ���ȴ�ή��������CIKϸ���ı��ͱ仯���ȡ�������ʵ��ָ��CIKϸ�����������Ʋ�����ҪIL-2���������������Zoll�ȵ��о��������������������IL-2��CIKϸ������ֳ��ɱ�˹����дٽ����á�Schmidt-Wolf��������IL-2����Ƭ�ε����������õ紩������CIKϸ������ת����ʵ����������תȾ��ϸ���ɷ��ڽϸ�ˮƽ��IL-2��330-1 800pg/106 cell/24h��ƽ����836pg/106 cell/24h ������ȻתȾǰ��CIKϸ���������Ĥ���ױ��ﲢ�������仯����������תȾ��CIK ϸ������ֳ�ʺ�ϸ��ɱ�˻����Ͼ�����δתȾϸ����
�Ʊ�����
��ȡ���ߵ�����Ѫ����37�棬5%�Ķ�����̼�������з���2h��ʹ�ø������Ļ����룬�ռ�����ϸ���������⣨ģ�������ڻ��������������ϸ��������CD3������IFN-R��IL-2��������ÿ��2-3�컻һ������Һ����7��11��13��15���ռ�CIKϸ����CD3+��CD56+ϸ��Ѹ�����࣬������ǰ�����ɴ�1000�����ϡ�
CIKϸ���Ʊ�����
CIKϸ���Ʊ�����
��չ����
1985���������ҽ���״η��ִ������IL-2��������Խ��ܰ�ϸ�������ɾ��к�ǿ����ɱ�����õ�ϸ������ΪLAKϸ�����Ժ���������Һ�м���CD3����¡������Խ���Щϸ��������ɱ�˻������ʮ����������������Ҳ�����ߣ�����ϸ����ΪCD3AK���������������������Һ�м���INF-r��IL-1��ϸ�����ӣ��õ�����CD3\CD8\CD56���Ե�����ϸ��Ⱥ����Щϸ����ΪCIK�����LAKϸ����ֳ�������࣬ɱ��������ǿ��
1991������˹̹����ѧSchmidt Wolf�����״α�����CIKϸ����
�ٴ�Ӧ�ñ༭
����1996���������CIKϸ�����Ƽ����ٴ�Ӧ�õ��о����ķ������������г����ټҵ�ҽ�Ƶ�λ��չ�˸������Ƽ���������ٴ�Ӧ�����о���
���ݹ��ʹ���ϸ�������ٴ�Ӧ�õĽ�չ״����2009��5��1���������䷢ִ�еġ�ҽ�Ƽ����ٴ�����Ӧ�ð취��������������ϸ�����Ƽ�������Ϊ����ҽ�Ƽ�����
�ΰ����߽�����������CIK���ƺ�1�ꡢ3�ꡢ5����������ڱȽϣ�������ͼ��
��Ӧ֢
CIKϸ���������ڹ���ϸ�������Ʒ� [3] ������CIKϸ�����������Ƿ�MHC�����Եģ�������������֯���͵����ƣ���˶��κ�һ����������ɱ�����ã����Ը߿�ԭ����İ�֢��Ч��ã��磺����Ѫ������ɫ��������ϸ������ת�����������Ǻνܽ����ܰ����ȡ�������֢��ΰ����᳦�������ٰ����ΰ���θ�������ȣ�CIKϸ���������нϺõ���Ч��CIKϸ�������������κ�һ�ڵİ�֢���ߣ������������������������Ż��ƺ��������ɽ�С�Ļ���Ч���á��������������Ż��ƻ���Ѫ��ϸ����ֲ��������С����������������ֹ��ϸ����ɢ��������������������������ٶ��Է�Ӧ�ȷ��������Ҫ���á�ijЩ���ʺ��������������ܷŻ��Ƶ��������������ߣ�CIKϸ�����ƿ�������������������ӳ���������ʱ�䡣
CIK ϸ��ͨ��ֱ��ɱ�ˡ����ڶ���ϸ�����ӵ�ֱ����������ϸ���������յ�����ϸ���������������á������ִ��ɱ�˹��ܵ�ϸ���ڻ��������ִ���书�ܣ���˥��Լ2����һ���¡������ϸ���а���һ���ּ���ϸ�����ɴ�������ʮ�꣬��������Ӧ�̼���Ѹ�������ڻ��ɱ�˰�ϸ��������CIKϸ�����Ƶ��Ƴ̼��ԭ����Ϊһ���£��������ȼ��һ���������������Ƴ̣��Ժ�ÿ������һ���Ƴ̡�Natural killer cells and autoimmunity.
French AR, Yokoyama WM - Arthritis research & therapy (2003)Affiliation: Howard Hughes Medical Institute, Division of Rheumatology, Department of Medicine, Washington University School of Medicine, St Louis, MO, USA. yokoyama@im.wustl.edu
Abstract
Autoimmune diseases are often characterized as clinical syndromes caused by the inappropriate activation of T or B cells resulting in systemic or organ-specific damage. However, studies support a role for the innate immune system, and in particular natural killer (NK) cells, in stimulating or suppressing autoimmunity. This review focuses on recent research elucidating a potential immunoregulatory role for NK cells in modulating T and B cell-mediated autoimmunity.
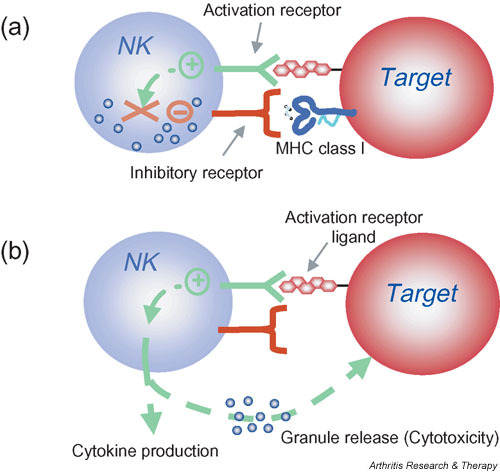
Natural killer (NK) cell activation is controlled by the integration of signals from activation and inhibitory receptors. (a) Inhibitory NK cell receptors recognize self MHC class I and restrain NK cell activation. (b) When unimpeded by the inhibitory receptors, binding of NK cell activation receptors to their ligands on target cells results in NK cell stimulation. In the absence or downregulation of self MHC class I on the target cells, these stimulatory signals are no longer suppressed, resulting in NK cell responses including cytokine production and granule release leading to cytotoxicity. Note that this model indicates that NK cells do not kill by default; that is, when MHC class I inhibition is absent, the NK cell must still be stimulated through activation receptors. Moreover, whether or not an individual NK cell is activated by a target is determined by this complex balance of receptors with opposing function and expression of the corresponding ligands. In general, however, inhibition dominates over activation. Finally, NK cells can be directly stimulated by cytokines such as interleukin-12 that trigger the production of other cytokines by NK cells (not shown). These direct cytokine-mediated responses are not affected by MHC class I expression.
© Copyright Policy
F1: Natural killer (NK) cell activation is controlled by the integration of signals from activation and inhibitory receptors. (a) Inhibitory NK cell receptors recognize self MHC class I and restrain NK cell activation. (b) When unimpeded by the inhibitory receptors, binding of NK cell activation receptors to their ligands on target cells results in NK cell stimulation. In the absence or downregulation of self MHC class I on the target cells, these stimulatory signals are no longer suppressed, resulting in NK cell responses including cytokine production and granule release leading to cytotoxicity. Note that this model indicates that NK cells do not kill by default; that is, when MHC class I inhibition is absent, the NK cell must still be stimulated through activation receptors. Moreover, whether or not an individual NK cell is activated by a target is determined by this complex balance of receptors with opposing function and expression of the corresponding ligands. In general, however, inhibition dominates over activation. Finally, NK cells can be directly stimulated by cytokines such as interleukin-12 that trigger the production of other cytokines by NK cells (not shown). These direct cytokine-mediated responses are not affected by MHC class I expression.Although NK cells are prepared to kill abnormal cells and rapidly release cytokines, they are normally restrained by inhibitory receptors that recognize target-cell-expressed MHC class I molecules and allow NK cells to survey tissues for normal MHC class I expression (Fig. 1). When MHC class I molecules are downregulated or absent, NK cells are released from the inhibitory influence of these receptors and kill target cells more efficiently ('missing self' hypothesis) [14]. It has been proposed that NK cells all express at least one inhibitory receptor that recognizes self MHC to provide NK cell tolerance and to prevent inappropriate NK cell responses directed at self [15,16]. However, release from inhibitory receptor effects does not automatically lead to NK cell activation against cellular targets. NK cells also express different combinations of various activation receptors, allowing them to respond to ligands on potential target cells [17-20]. The ligands for activation receptors are often closely related to the ligands for inhibitory receptors. In other cases, the activation receptor ligands may be upregulated in response to stress or infection, as illustrated by the upregulation of the NKG2D ligand MHC class I-related chain A (MICA) during infection [21-23].
Natural killer (NK) cell activation is controlled by th | Open-i
https://openi.nlm.nih.gov/detailedresult?img=PMC400423_ar1034-1&req=4��
Immunology. 2007 Jun; 121(2): 197�C206.
Interleukin-12- and interferon-��-mediated natural killer cell activation by Agaricus blazei Murill
Eri Yuminamochi,1,2,* Taisuke Koike,1,2,* Kazuyoshi Takeda,1 Isao Horiuchi,2 and Ko Okumura1
1Department of Immunology, Juntendo University School of Medicine, Tokyo, Japan
2Japan Applied Microbiology Research Institute Ltd, Yamanashi, Japan
ABSTRACT
Dried fruiting bodies of Agaricus blazei Murill (A. blazei) and its extracts have generally used as complementary and alternative medicines (CAMs).Here, we report that the oral administration of A. blazei augmented cytotoxicity of natural killer (NK) cells in wild-type (WT) C57BL/6, C3H/HeJ, and BALB/c mice.
Augmented cytotoxicity was demonstrated by purified NK cells from treated wild-type (WT) and RAG-2-deficient mice, but not from interferon-�� (IFN-��) deficient mice. NK cell activation and IFN-�� production was also observed in vitro when dendritic cell (DC)-rich splenocytes of WT mice were coincubation with an extract of A. blazei.
Both parameters were largely inhibited by neutralizing anti-interleukin-12 (IL-12) monoclonal antibody (mAb) and completely inhibited when anti-IL-12 mAb and anti-IL-18 mAb were used in combination. An aqueous extract of the hemicellulase-digested compound of A. blazei particle; (ABPC) induced IFN-�� production more effectively, and this was completely inhibited by anti-IL-12 mAb alone.
NK cell cytotoxicty was augmented with the same extracts, again in an IL-12 and IFN-��-dependent manner. These results clearly demonstrated that A. blazei and ABPC augmented NK cell activation through IL-12-mediated IFN-�� production.
Keywords: Agaricus blazei, NK cells, IFN-��, IL-12, cytotoxicityInterleukin-12- and interferon-��-mediated natural killer cell activation by Agaricus blazei Murill
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2265935/��
Annu Rev Immunol. 1995;13:251-76.
Interleukin-12: a proinflammatory cytokine with immunoregulatory functions that bridge innate resistance and antigen-specific adaptive immunity.
Trinchieri G1.
Wistar Institute of Anatomy and Biology, Philadelphia, Pennsylvania 19104-4268, USA.
Abstract
Interleukin-12 (IL-12) is a heterodimeric cytokine produced mostly by phagocytic cells in response to bacteria, bacterial products, and intracellular parasites, and to some degree by B lymphocytes.IL-12 induces cytokine production, primarily of IFN-gamma, from NK and T cells, acts as a growth factor for activated NK and T cells, enhances the cytotoxic activity of NK cells, and favors cytotoxic T lymphocyte generation.
In vivo IL-12 acts primarily at three stages during the innate resistance/adaptive immune response to infection:
1. Early in the infection, IL-12 is produced and induces production from NK and T cells of IFN-gamma, which contributes to phagocytic cell activation and inflammation;
2. IL-12 and IL-12-induced IFN-gamma favor Th1 cell differentiation by priming CD4+ T cells for high IFN-gamma production; and
3. IL-12 contributes to optimal IFN-gamma production and to proliferation of differentiated Th1 cells in response to antigen. The early preference expressed in the immune response depends on the balance between IL-12, which favors Th1 responses, and IL-4, which favors Th2 responses.
Thus, IL-12 represents a functional bridge between the early nonspecific innate resistance and the subsequent antigen-specific adaptive immunity.
PMID: 7612223 DOI: 10.1146/annurev.iy.13.040195.001343Interleukin-12: a proinflammatory cytokine with immunoregulatory functions that bridge innate resistance and antigen-specific adaptive immunity. - PubMed - NCBI
https://www.ncbi.nlm.nih.gov/pubmed/7612223��
��
Revisiting Interleukin-12 as a Cancer Immunotherapy Agent | Clinical Cancer Research
https://clincancerres.aacrjournals.org/content/24/12/2716��
Oxidant-induced cell death in lymphocytes �C mechanisms of induction and resistance
Fredrik B Thor��n
University of Gothenburg, Germany
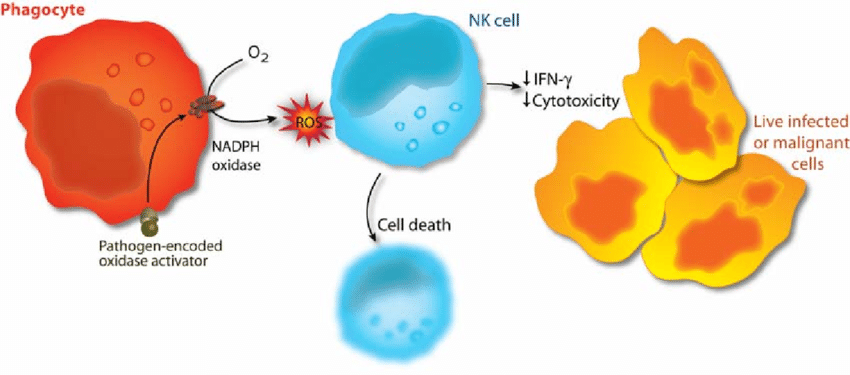
Reactive oxygen species (oxidants, oxygen radicals) produced by the phagocytic NADPH oxidase have pivotal roles in immunity. Patients lacking a functional NADPH oxidase suffer from chronic granulomatous disease, which is characterized by recurring bacterial infections and thus manifesting the importance of reactive oxygen species in host defense against bacteria.However, NADPH oxidase-derived radicals also efficiently inhibit lymphocyte-mediated immunity. Oxidant-induced inactivation of lymphocytes is reportedly a control mechanism for autoreactive lymphocytes and hence prevents autoimmunity. In malignant diseases, oxygen radicals have been proposed to contribute to the characteristic state of anergy of cytotoxic lymphocytes, which prevents immune-mediated rejection of the tumor.
Studies of the mechanisms of radical-induced inactivation of lymphocytes may therefore be helpful in understanding the pathophysiology of important disease entities. The first paper in this thesis shows that oxidant-induced functional inhibition and cell death in cytotoxic lymphocytes is critically dependent on cooperation between a nuclear enzyme involved in DNA repair, PARP-1, and a mitochondrion-derived protein, AIF.
The results presented in Paper II demonstrate that pharmacological inhibition of the PARP-1 enzyme not only prevents oxidant-induced cell death, but also preserves functions of cytotoxic lymphocytes, such as cytotoxicity against malignant cells, cytokine production, and proliferation.
Paper III shows that subsets of natural killer (NK) cells display differential sensitivity to oxygen radicals: the cytotoxic CD56dimCD16+ NK cells were found to be highly sensitive to oxidative inactivation and apoptosis, while the immunoregulatory, cytokine-producing CD56brightCD16- NK cells were highly resistant to the toxicity of oxidants.
These data were extended in Paper IV, in which the effect of oxygen radical-producing phagocytes on the expression of the activating NK cell receptors, NKp46 and NKG2D, was investigated. The expression of both receptors was efficiently downregulated on CD56dim NK cells, while the expression remained intact on CD56bright cells.
Recent data imply that reciprocal interactions between NK cells and dendritic cells (DCs) are important for the development of adaptive immunity. The results presented in Paper V demonstrate that DCs are equipped with an antioxidative system that efficiently protects cytotoxic cells from oxidant-induced inactivation.
��
Phagocyte activation as an immunoevasive strategy. In malignant and infected tissues, phagocytes are recruited with ensuing production of oxygen radicals. The release of oxygen radicals from phagocytes inactivates NK cells and other cytotoxic lymphocytes, enabling survival of malignant or virus-infected cells (adapted from 298).
��
��
��
DNA repair responses to PARP1
University of Texus, MD Anderson Canver Center
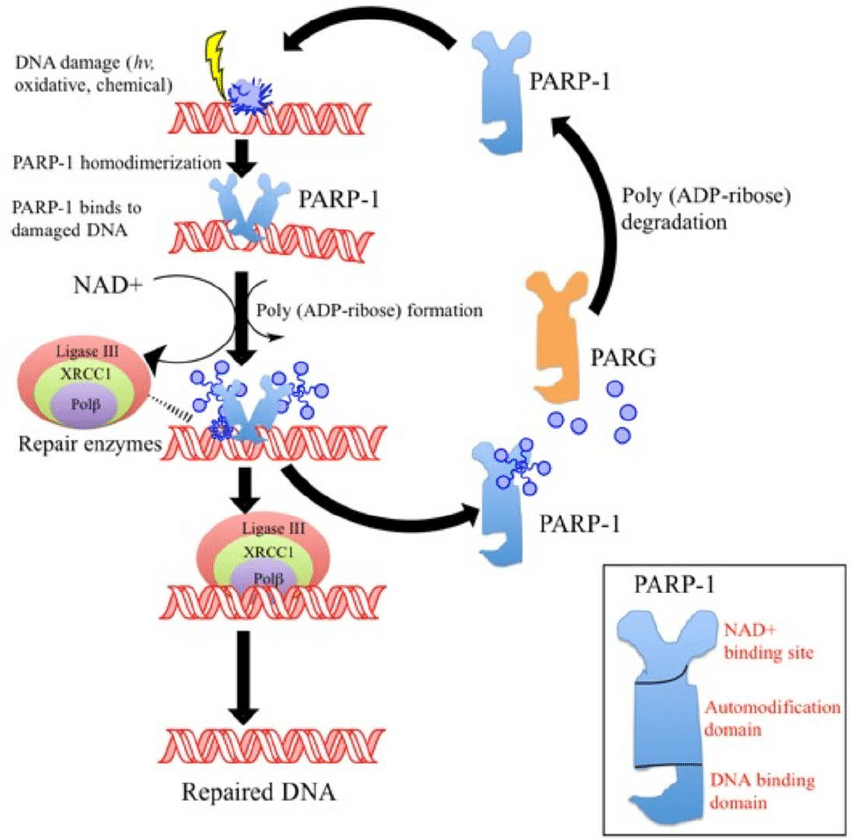
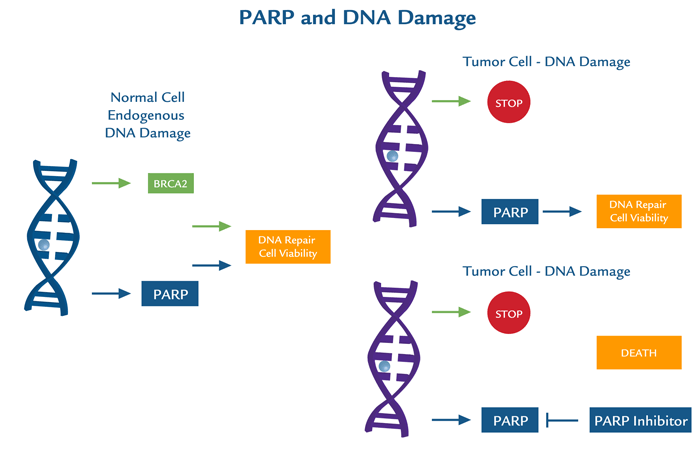
PARP1 encompasses a broad scope of DNA repair responses which makes it an attractive candidate for therapeutic inhibition, either alone or in combination with cancer chemotherapy or radiotherapy. Inhibition of PARP1 has potential for use in cancer treatment particularly by increasing tumor sensitivity to chemotherapeutic agents that damage DNA and by inducing synthetic lethality in tumor cells that are highly dependent on PARP1 due to deficiencies in DNA repair proteins (e.g., BRCA1/2).PARP1 and DNA damage
Other mechanisms by which PARP exerts its effects are through rapid mitochondrial dysfunction with membrane permeability transition, NAD+ depletion, and translocation of apoptosis inducing factor (AIF) from the mitochondria to the nucleus. Inhibition/manipulation of other components of the PARP pathway, including PAR and PARG may therefore, be valuable therapeutic interventions not only for cancer, but for other disease states.
MDAnderson - PARP1 DNA ToolkitIn collaboration with MD Anderson Cancer Center, a highly specific toolkit has been developed and validated to analyse PARP1 in a panel of control and siRNA knockdown cell lysates by multiple immunoassays. This toolkit can specifically and quantitatively distinguish PARP1 from 17 other PARP family members using highly specific antibodies and optimised methods.
Most clinically used PARP inhibitors bind to conserved regions in the PARP domains that permits cross-selectivity with other PARPs containing homologous catalytic domains. Thus, the differences between therapeutic effects and unwanted side effects resulting from pan-PARP inhibition compared to selective inhibition are not well understood. The development and use of more selective agents targeting other domains that make up PARPs, will help answer important questions concerning PARP inhibitors as chemotherapy. A clear understanding of the inhibition profiles of PARP inhibitors will not only enhance the understanding of the biology of individual PARPs, but may provide improved therapeutic options for patients.
Do not hesitate to contact us to test this new toolkit!https://www.tebu-bio.com/blog/2015/10/13/dna-repair-responses-to-parp1/
��

.jpg)