��
����ϸ������ - ����ϸ�����ά����C����ά����Cɱ���Ļ���
Stromal Cell Oxidation-A Mechanism by Which Tumors Obtain Vitamin C and killed by Vitamin C
Department of Medicine, Memorial Sloan-Kettering Cancer Center, New York, New York 10021
ͼʾ4�������ά����C������ϸ����TUMOR CELL) �е�ת�˺ͻ��ۻ��ơ�����Ѫ�ᣨAscorbic Acid) ��ѪҺ��ά����C����Ҫ��ʽ����������������ͨ������������(Superoxide Anion) �����������⿹��Ѫ�ᣨDHA)����ˣ�����(Oxidation)������ת��ά����C�ĵ��ڲ��衣Ȼ�����⿹��Ѫ��ͨ������ϸ�������GLUT1ת�ˡ�Ȼ������ϸ���ڵ����⿹��Ѫ��(DHA)����ԭΪ����Ѫ�ᣨAscorbic Acid)������������ͨ��GLUT1ת�ˣ���˿���Ѫ�ᱻ����������ϸ���С�
ժҪ
�����������ܺ��и�Ũ�ȵĿ���Ѫ�ᣬ�����Ƕ�������λ��ά����֪֮���١�ijЩ�ػ�ϸ������ֱ��ͨ������Ѫ����Эͬת�˵��ף�SVCT)���俹��Ѫ�ᣬ���ڴ����ϸ���У�ά����C�����⿹��Ѫ�����ʽͨ���ٽ�������ת�˵��ף�GLUT�����룬Ȼ����ϸ���ڻ�ԭ������Ϊ����Ѫ�ᡣ
�о����ѽ�������Ѫϸ������Ƥϸ��������ֲ���С��ע�俹��Ѫ������⿹��Ѫ��Ļ��ۡ��������Ѫϸ������Ƥ����ϸ��ϵֻ������������������ʽ��ά����C�����⿹��Ѫ�ᣬDHA��;Ȼ��������С������Ϊ������ֲ������ʱ��������ʩ�÷����Ա�ǵĿ���Ѫ���Ѹ�ٻ���ά����C.ͨ����D-�����ǣ�D-glucose)������L-�����ǵľ���������֤����GLUT����������ֲ������ά����C����ȡ��
���ڶ���ϸ������ֱ��ת�˿���Ѫ�ᣬ�����ƶϿ���Ѫ�������������б����������⿹��Ѫ��(DHA)��ע�俹��Ѫ��Ķ�����ά����C����������ͨ����ͬʩ�ó��������绯ø(superoxide dismutase)�������ƣ�����ζ�ų����������ڿ���Ѫ��������������á�
������Ƥ��ϸ��ϵ���������в��ܲ������������ӣ����������������ֲ����ȷʵ���ڡ����ǵ��о�������GLUTs�����⿹��Ѫ���ת�����������ά����C��һ���ֶΣ����������������е�ϸ�������ij�����������������Ѫ����Ϊ������������ʽ��ά���صĻ��ơ���
����
��ʳ�е�ά����C������������鳤�ද��������Ҫ����Ϊ���Dz�����������������һ���ڸ���ϳ�����ά���أ�1������Ȼ���Ƕ�ά����C��ά����Cȱ��֢���˽�ܶ࣬�����ڰ�֢��ά���ص�����ѧ֪ʶȴ���٣�2��������ά����C��ά���������߹��̺�������������ij��֤�ݣ������ձ���Ϊ����ά����C������ǿ������ϵͳ��3,4��������ά����C�İ�֢����ͨ����Ϊ��������ǿ�Կ���֢�����߷�������Щ������ٹ�ע��֢������Ӫ������ϸ��������������ȡά����C��5,6���������о����������е�ά����CŨ�ȸ����ڽ���������֯��7����Ȼ���������������ۻ�ά����C�Ļ����в������
������ǰ�������⿹��Ѫ�ᣬ������ʽ��ά����C��������ͨ���ٽ���������ת�˵��ף�GLUTs�� 3ת�ˣ�8��������צ���ĸϸ����GLUT1��GLUT2��GLUT4�ı��︳������ȡ���⿹��Ѫ������������⿹��Ѫ���ڻ�ԭΪ����Ѫ�����ϸ���ڱ��������ǻ�ȷ��GLUT����������������ϸ����������Ѫ��ϸ��ϵHL60������ͻ���ά����C��8,9,10��������Щϸ���У����⿹��Ѫ��ͨ��ϸ��Ĥת�˲��Ի�ԭ��ʽ����Ѫ����ۣ�����Ѫ���ͨ��GLUT˫��ת�ˣ�8,9,10���� GLUT1��Ѫ������������ת�����⿹��Ѫ��Ҳ�Ǵ��Ի��ά����C�Ļ��ƣ�11��������Ѫ�ᣬ��ѪҺ��ά����C����ʽ�����״���Ѫ�����ϣ������⿹��Ѫ�ᴩ��Ѫ�����ϲ���Ϊ����Ѫ����������С�
����Ѫ��Ҳ����ֱ��ͨ��һϵ��Na +�����Կ���Ѫ��ת�˵��ף�SVCT)ת�ˣ���Щת�˵�������ѱ����Ӽ�����12��������δ����WBC�л��о��а�����ǰ���٣����ٺ���Ѫ����ϸ��ϵ��Na +�����Կ���Ѫ����ȡ��֤�ݣ�9,10����
���о�����ǰ���٣����ٺ���Ѫ����������ֲ����ͨ��GLUT���������ʽ��ά����C.���������������������ӣ�������Ѫ�����������⿹��Ѫ�ᣬ����ά����C�Ŀ�������ʽ����������⿹��Ѫ��ͨ����ԭ�ɲ���ת����ʽ��ά����C��������Ѫ����������������С�
��
���Ϻͷ���
�����о�
��
���������Ұ�֢�о��� - ������˰�֢���Ļ��4��6���������������BALB / c����С��Ƥ��ע�䡣������Ũ������ϸ������ู��HL-60ϸ����2.0��107ϸ��; LNCaPϸ����3.0��106ϸ��; MDA468ϸ����1.0��107ϸ��; ���ֺ�3~4�ܣ��ɲⶨ~1.0��1.0��1.0cm��������CEMϸ����2.0��107�����ؽ��Ļ���Ĥ��Matrigel; Collaborative Research��Bedford��MA����1��1�������ע��LNCaPϸ������ѭ�����о�����ȷ���˵���ʹ�ö�����ƶ���
��֯ת�˺ͻ����о�
Я��������ֲ������С����5��Ci��1- [1-14C]����Ѫ�ᣨ�Ȼ��ԣ�6.6mCi / mmol; DuPont-NEN��Boston��MA����[14C]���⿹��Ѫ���[����-1-3H]ע��β���������ǣ��Ȼ�ȣ�20.0Ci / mmol; DuPont-NEN���� [14C]������ʵ����ͨ����[14 C]����Ѫ���뿹��Ѫ������ø��ÿ1.0mmol L-����Ѫ����1��λ����Դ���Ϲ�������; Sigma Chemical Co.��St.Louis��MO��һ��������⿹��Ѫ�ᡣ��DTT��0.1mmol / l; Sigma�����뵽ά����C�Ƽ��С�Ȼ���г����ٺ���������70���״����Ƚ�����ǰ������11��������Ʒ������˸���ײⶨ��HPLC���ڼ״������Ͻ���HPLC������1mmol /��EDTA����Whatmanǿ�����ӽ���Partisil 10 SAX��4.6��25cm������Whatman��Inc.��Clifton��NJ���Ϸ���HPLC��Ʒ��ʹ��Whatman��WCS�ܼ�������������System GoldҺ��ɫ���ǣ�Beckman Instruments��Inc.��Fullerton��CA�����ϴ��Һ�����ж��������м�����ͷ�����ͬλ�ؼ�����������á�ͨ��265nm��������Ⱥͷ����Լ�⿹��Ѫ�ᡣ���⿹��Ѫ����265nm��û����ʾ����ȣ���˽�ͨ�������Լ�⡣��2-����-D-�����ǣ�Sigma����L-�����ǣ�Sigma����SOD����������������ԭø�������˺�ϸ��; Sigma����������ø��H2O2������ԭø������С�����; Sigma�����й�ע��ʵ�顣
�������������ɵIJ�����
�������������Ӳ���Ϊ��SOD�����Ƶ�ϸ��ɫ��C��ԭ�� ��ϸ��ϵ�����ʷ���������������37������HBSSԤ����15���ӣ��ú���ϸ��ɫ��C��HBSSϴ��һ�Σ����뺬�л�SOD��200��λ/ ml����1mg / mlϸ��ɫ��C��HBSS�з����� ҡ���� ��PMA��Sigma����80nm����Ũ��������ϸ��ϵ�� ��60����ʱ����ϸ����ȡ�������������ڱ�����ֹͣ��Ӧ��������ȡ550nm��������ȡ� ϸ��ɫ��C�ij������������Ի�ԭ��ʾΪʹ��21.1nm-1��cm-1������ϵ�������л�û��SOD������·�����ϸ��֮�������Ȳ��졣����������ϸ�������������������
���
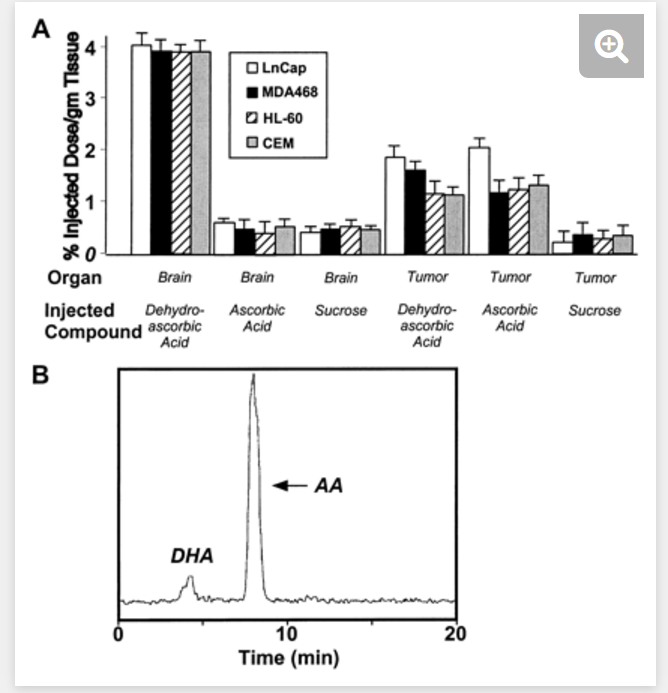
������������ֲ������С����[14 C]����Ѫ�ᣬ[14 C]���⿹��Ѫ���[3H]����ע�䵽β�����У�����ע���1���Ӵ�������1���Ӻ���������ֲ������з��ִ�Լ4����ע�����⿹��Ѫ������ԣ���ʾΪÿ����֯��ID�İٷֱȣ���ͼ1A���������������ǰ���о�һ�£�11����ע��Ŀ���Ѫ���������1����ʱ�������Ƚ��в����������ԣ�֤ʵ����Ѫ�����ͨ��Ѫ�����ϡ����Dz�����л�����䣬�����������Ѫ������ı���13����������ֲ������1����ʱ�ۻ�ע�����⿹��Ѫ�ᣬLNCaP��ǰ���٣������е�������֯Ϊ〜2.0��ID / g��MDA468�����٣�����Ϊ1.6��ID / g��HL-60Ϊ1.2��ID / g����ϸ����Ѫ���� ��CEM�е�������1.2��ID / g��T-�ܰ�ϸ����Ѫ��;ͼ1A⇓��������δ��ת�˵������С�����Բ�ͬ����������Ѫ��ע�붯��ʱ��������ֲ�����������շ����ԡ� LNCaP�����ۻ�〜2.3��ID / g��MDA-468 1.2��ID / g��HL-60 1.3��ID / g����CEM 1.3��ID / g������֯��ͼ1A��⇓����������Я��������ֲ���С��������Ƚ��ļ״����ֵ�HPLC�������Լ����ۻ���ά����C����ʽ�������������ע�����⿹��Ѫ��Ķ����ע�俹��Ѫ��Ķ����У��������ۻ���ά����C��ע�����⿹��Ѫ��Ķ����ע�俹��Ѫ��Ķ���> 86������Ѫ�ᣨͼ1B��⇓��
ά����C�������ϸ����������ϸ������������ʽ��Ϊ���⿹��Ѫ��ͨ���ٽ���������ת�˵��ף�GLUTs;�ο�����8,9,10����ȡ����ˣ�����Ѫ�������ϸ����Χ�����б�����Ϊ���⿹��Ѫ�ᣬ��ͨ��GLUTת�˵�ϸ���У����������ԭ���Կ���Ѫ�����ʽ��������Ϊ���������ϸ���������俹��Ѫ�ᣬ���Ǽ���ע��Ŀ���Ѫ�������������б����������⿹��Ѫ�ᣬȻ��ͨ��GLUT���䡣Ϊ��ȷ������Ѫ���Ƿ������ַ�ʽ��������ϸ����������D-��L-�����ǽ�������ʵ�飬��ΪGLUTѡ���Ե�ת��D-�����Ƕ�����L-�����ǡ�����ʾD-���������Ǻ�D-�������Լ��������ķ�ʽͨ��GLUT�����������⿹��Ѫ�����ȡ�ߴ�70����11�����ں���С���У�D-�����������������⿹��Ѫ��Ϳ���Ѫ���ת�˺ͻ��ۣ�ͼ2��A-D��⇓�� 5��ʱ��������о�ת�ˣ�����ά����C��ȡ�ļ������������ƴ�ԼΪ35-50������ע�����⿹��Ѫ��Ϳ���Ѫ�ᣩ�� 1���Ӻ�5���ӵ�ʱ����������ͻ��ۣ���֤����ά����C��ȡ��������������40-65��������ƣ���ע�����⿹��Ѫ��Ϳ���Ѫ�ᣩ������L-�����Ƕ�������ֲ����ά����C��ת�˻����û��Ӱ�죨����δ��ʾ������ע�俹��Ѫ���С���У����������Ƕ�����ά����C��ȡ�ͻ��۵����Ʊ�������Ѫ��ת��Ϊ���⿹��Ѫ�ᣬ�Ӷ�ͨ��GLUTת�˵�����ϸ���С�
���Ǽ��迹��Ѫ�������������б����������������� Ϊ�˲������������ǹ�ͬע��Я������Ѫ���SOD����������ø����ˮ��������ֲ��Ķ�� ����SOD�ͷ����Ա�ǵĿ���Ѫ��Ķ�������ά����C���ۼ���Լ50�����������⿹��Ѫ���SOD��ͬע��Ķ�����ά����C����������û�б仯��ͼ3A��⇓�� �Կ���Ѫ���������ø�Ĺ�ͬ��ҩû��Ӱ�죬����������������ڽ�����Ѫ�����������⿹��Ѫ�᷽��û�����ã�ͼ3A����
���Dz���������ϸ�������������������ӵ������� ���������ӵIJ�������HL-60ϸ��ϵ֤����ͼ3B������������ϸ��ϵ�����в������������ӵ������� �������ϣ�����PMA�ʱ��HL-60ϸ��ϵ���ӳ�������IJ�����������ϸ��ϵ��ʾû��PMA�յ��ij��������Ӳ��������ױ�HL-60ϸ��ϵ��ȣ���ʹ�������HL-60������ֲ�����ij��������Ӳ���Ҳ������3���� ��Ϊ��ϸ��ϵ��ͬ�������������ֲ�������в������������ӵ��������������ǵó����ۣ����������еķ�����ϸ���Dz������������ԭ��
��
����
������ͼͨ��������������С����ʹ��������������ֲģ�Ͳ�����[14C]����Ѫ�����ȡ��ȷ��������ϸ�������������ȡά����C. ע�俹��Ѫ�������Ѹ��ռ���˱�ǩ����������֮ǰ�Ѿ�������֤��ϸ������ֱ��ת�˿���Ѫ�ᣨ8,10���������Ʋ�����ϸ�������⿹��Ѫ�����ʽ��ȡά����C. �۲쵽��ע��������������ȡ������֤ʵ�������Ĺ۵㣬��ϸ�������⿹��Ѫ�����ʽ���ͨ���ٽ���GLUT�����ά����C .��ΪGLUT������������ʽ��ά����C�����⿹��Ѫ�ᣬ������Ϊϸ��û��������ȡά����C�Ļ��ƣ����Ժ����Կ���Ѫ�������������б����������⿹��Ѫ�ᡣ����ͬʱȱ��14C��������ȡ��������Ѫ����ѭ����δ��������ͨ��SOD����ע�俹��Ѫ��Ķ�����ά����C����ȡָ���˳����������ڿ���Ѫ�������е����á���Ƥ����ȱ�����������ӵIJ�����������ֲ�����г��������ӵIJ�������������ϸ���dz��������ӵ���Դ����ˣ������������Ѫ��������������ͨ���Ƕ��Ի���ϸ���������������Ӷ����������Ѿ�ͨ������NADPH����ø�ij���άϸ������������ϸ��������ϸ������Ƥϸ��֤ʵ������ͳ��������ӵIJ�����14,15,16,17��������ʾ��������ϸ����HL-60ϸ���Գ������������Է�ʽ����ϸ���⿹��Ѫ�ᣨ16,18,19����ijЩ���������磬������Ѫ�������о��е�HL-60������ֲ���ֱ����������Ѫ�ᣬ��������������Ƥ����������������������ϸ����ʵ�ָù��ܡ�ͼ4�ܽ�������ת�˺ͻ���ά����C��ģ�͡�
����Ѫ����Эͬת�˵��ף�SVCT�����������������У�12�����������������о���δ����������ϸ��������ϸ����HL-60ϸ����T�ܰ�ϸ��ϸ��ϵ��ǰ���ٰ������ٰ�ϸ��ϵ�е��������Կ���Ѫ����ȡ ��SVCT)��9,10������ˣ�ͨ��GLUT�����⿹��Ѫ�����ʽ��ȡά����C�ƺ���ά����C��ȡ��һ����ơ���������֮ǰ��ʾ������Ѫ����Эͬת�˵�������ijЩ����ά����C��ȡ�������ã�6����
ά����C������ϸ���е�ȷ�й����в�������о���������������֯��ȣ�ijЩ����������ά����C��Ũ�����ӣ�7����ά����C������ijЩ����ϸ������������Ҫ��20����ά����C���������ӻ����Ѫ�忹��Ѫ��ˮƽ��21������˿��ܻ�����ά����C������Ũ�ȡ�ϸ����ά����CŨ�ȵ����ӿ��ܶ����������������ķ�Ӧ��������Ӱ�졣����Ӧ���뻯�ƺͷ��������йء���������ά����C������ڰ�֢�����е����õ��о�ͨ��������������������˷���û���κ��洦��22�������в�������������и�Ũ�ȵ�ά����C�Ƿ����ṩ���д�л���ƵĶ���ϸ����
��
https://s.click.taobao.com/MuACULw
https://s.click.taobao.com/U7CCULw
��
Stromal Cell Oxidation-A Mechanism by Which Tumors Obtain Vitamin C and killed by Vitamin C
A proposed mechanism of vitamin C transport and accumulation in tumors. Ascorbic acid, the predominant form of vitamin C in blood, is oxidized to dehydroascorbic acid in the tumor microenvironment by superoxide anion. Oxidation is, thus, a regulatory step for tumor transport of vitamin C. The dehydroascorbic acid is then transported through GLUT1 at the surface of the tumor cell. The dehydroascorbic acid in the tumor is then reduced to ascorbic acid, which is trapped in the tumor because it cannot be transported through GLUT1.
��
David B. Agus, Juan C. Vera and David W. Golde
Abstract
Human tumors may contain high concentrations of ascorbic acid, but little is known about how they acquire the vitamin. Certain specialized cells can transport ascorbic acid directly through a sodium ascorbate cotransporter(SVCT), but in most cells, vitamin C enters through the facilitative glucose transporters (GLUTs) in the form of dehydroascorbic acid, which is then reduced intracellularly and retained as ascorbic acid.
Mice with established hematopoietic and epithelial cell xenografts were studied for the accumulation of injected ascorbic acid and dehydroascorbic acid. Most hematopoietic and epithelial tumor cell lines can only transport vitamin C in the oxidized form (dehydroascorbic acid) in vitro; however, when grown as xenografts in mice, they rapidly accumulated vitamin C after administration of radiolabeled ascorbic acid. The involvement of the GLUTs in vitamin C uptake by the xenografted tumors was demonstrated by competitive inhibition with d-glucose but not l-glucose.
��
Because the malignant cells were not capable of directly transporting ascorbic acid, we reasoned that the ascorbic acid was oxidized to dehydroascorbic acid in the tumor microenvironment. Tumor accumulation of vitamin C in animals injected with ascorbic acid was inhibited by coadministration of superoxide dismutase, implying a role for superoxide anion in the oxidation of ascorbic acid.
��
Whereas the epithelial cancer cell lines could not generate superoxide anion in culture, the minced xenograft tumors did. Our studies show the transport of dehydroascorbic acid by GLUTs is a means by which tumors acquire vitamin C and indicate the oxidation of ascorbic acid by superoxide anion produced by cells in the tumor stroma as a mechanism for generating the transportable form of the vitamin.
Introduction
Dietary vitamin C is critical for humans and other primates because we cannot synthesize the vitamin in the liver as do most other animals (1) . Whereas much is known about vitamin C and vitamin C deficiency states, there is little information regarding the physiology of the vitamin in cancer (2) . Given the well-documented role of vitamin C in the maintenance of normal immune processes and host defense, it is popularly believed that supplemental vitamin C ��strengthens�� the immune system (3 , 4) . Patients with cancer who take vitamin C generally believe that it can enhance immune defense against the cancer. These notions give little attention to the nutritional needs of the cancer itself. Cancer cells readily take up vitamin C in vitro (5 , 6) , and studies have demonstrated high vitamin C concentrations in neoplasms compared with the adjacent normal tissue (7) . The mechanism by which cancers accumulate vitamin C in vivo, however, is unknown.
We previously found that dehydroascorbic acid, the oxidized form of vitamin C, is transported in vitro through the facilitative GLUTs 3 (8) . Expression of GLUT1, GLUT2, and GLUT4 in Xenopus oocytes conferred the ability to take up dehydroascorbic acid which was retained intracellularly after reduction to ascorbic acid. We also established that GLUTs are involved in the transport and accumulation of vitamin C by normal human neutrophils and the myeloid leukemia cell line, HL60 (8, 9, 10) . In these cells, dehydroascorbic acid is transported across the cell membrane and accumulated in the reduced form, ascorbic acid, which is not transportable through the bidirectional GLUTs (8, 9, 10) . The in vivo transport of dehydroascorbic acid by GLUT1 at the blood-brain barrier is also a mechanism by which the brain acquires vitamin C (11) . Ascorbic acid, the form of vitamin C in the bloodstream, does not readily cross the blood-brain barrier, whereas dehydroascorbic acid crosses the blood-brain barrier and accumulates in the brain as ascorbic acid.
Ascorbic acid may also be transported directly through a family of Na+-dependent ascorbic acid transporters that have recently been molecularly characterized (12) . We have found no evidence of Na+-dependent ascorbic acid uptake in WBCs or in the prostate, breast, and hematopoietic tumor cell lines included in this study (9 , 10) .
This study demonstrates that prostate, breast, and hematopoietic human xenograft tumors obtain vitamin C in the oxidized form through the GLUTs. The tumor microenvironment generates superoxide anions that oxidize ascorbic acid to dehydroascorbic acid, the transportable form of the vitamin. The transported dehydroascorbic acid is trapped in the tumors by reduction to the nontransportable form of vitamin C, ascorbic acid.
Materials and Methods
Animal Studies.
Four- to 6-week old nude athymic BALB/c male mice were obtained from the National Cancer Institute-Frederick Cancer Center and injected s.c. into the flank with tumor cells at the following concentrations: HL-60 cells, 2.0 �� 107 cells; LNCaP cells, 3.0 �� 106 cells; MDA468 cells, 1.0 �� 107 cells; and CEM cells, 2.0 �� 107. Three to 4 weeks after inoculation, ∼1.0 �� 1.0 �� 1.0 cm tumors could be measured. The LNCaP cells were injected in a 1:1 volume ratio with reconstituted basement membrane (Matrigel; Collaborative Research, Bedford, MA). Institutional guidelines for the proper and humane use of animals in research were followed.
Tissue Transport and Accumulation Studies.
Mice bearing xenograft tumors were injected into the tail vein with 5 ��Ci of l-[1-14C]ascorbic acid (specific activity, 6.6 mCi/mmol; DuPont-NEN, Boston, MA), [14C]dehydroascorbic acid, or [fructose-1-3H]sucrose (specific activity, 20.0 Ci/mmol; DuPont-NEN). [14C]Dehydroascorbic acid was generated in all experiments by incubating [14C]ascorbic acid with ascorbate oxidase (1 unit per 1.0 mmol of l-ascorbate, derived from Cucurbita species; Sigma Chemical Co., St. Louis, MO). DTT (0.1 mmol/liter; Sigma) was added to the vitamin C preparations. The organs and tumors were then dissected out and homogenized in 70% methanol. Samples were processed for scintillation spectrometry or HPLC as described previously (11) . HPLC was performed on the methanol fraction with the addition of 1 mmol/liter EDTA. HPLC samples were separated on a Whatman strong anion exchange Partisil 10 SAX (4.6 �� 25 cm) column (Whatman, Inc., Clifton, NJ). A Whatman-type WCS solvent-conditioning column was used, and the eluates were monitored with a System Gold liquid chromatograph (Beckman Instruments, Inc., Fullerton, CA) with a diode array detector and radioisotope detector arranged in series. Ascorbic acid was monitored by absorbance at 265 nm and by radioactivity. Dehydroascorbic acid shows no absorbance at 265 nm, and so it was monitored only by radioactivity. Coinjection experiments were performed with 2-deoxy-d-glucose (Sigma), l-glucose (Sigma), SOD (superoxide oxidoreductase, from human erythrocytes; Sigma), and catalase (H2O2 oxidoreductase, from mouse liver; Sigma).
Measurement of Superoxide Anion Production.
Superoxide anion production was measured as the SOD-inhibitable reduction of cytochrome c. Cell lines and freshly isolated minced tumors were preincubated with HBSS for 15 min at 37��C, washed once with HBSS containing cytochrome c, and incubated with 1 mg/ml cytochrome c with or without SOD (200 units/ml) in HBSS on a shaking table. PMA (Sigma) was added to the cell lines at a final concentration of 80 nm. At 60 min, the medium was removed from the cells and placed on ice to stop the reaction, and the absorbance at 550 nm was immediately read. Superoxide-specific reduction of cytochrome c was expressed as the difference in absorbance between cells incubated with or without SOD by use of an extinction coefficient of 21.1 nm−1��cm−1. Cell counts of minced tumor were derived from volumetric measurements.
Results
Mice with xenograft tumors were injected into the tail vein with [14C]ascorbic acid, [14C]dehydroascorbic acid, or [3H]sucrose and sacrificed 1 min after injection. Approximately 4% of the injected dehydroascorbic acid radioactivity (expressed as percentage of ID per gram of tissue) was found in the brain of the xenograft groups after 1 min (Fig. 1A) ⇓ , a result consistent with our previous work (11) . Injected ascorbic acid and sucrose yielded only trace radioactivity in the brain homogenate at 1 min, confirming that ascorbic acid did not readily pass the blood-brain barrier. Sucrose is not metabolized or transported, and therefore it is used as a marker of plasma volume (13) . The xenograft tumors accumulated injected dehydroascorbic acid at 1 min with ∼2.0%ID/g tumor tissue in LNCaP (prostate) tumors, 1.6%ID/g in MDA468 (breast) tumors, 1.2%ID/g in HL-60 (myeloid leukemia) tumors, and 1.2%ID/g in CEM (T-lymphocyte leukemia; Fig. 1A ⇓ ). Sucrose was not transported into the tumors. Distinct from the brain, the xenograft tumors readily took up radioactivity when ascorbic acid was injected into the animals. LNCaP tumors accumulated ∼2.3%ID/g, MDA-468 1.2%ID/g, HL-60 1.3%ID/g, and CEM 1.3%ID/g tumor tissue (Fig. 1A) ⇓ . HPLC analysis of the methanol fraction of the tumor homogenate from the xenograft-bearing mice was performed to identify the form of vitamin C accumulated. The results show that the vitamin C accumulated in the tumors was >86% ascorbic acid in animals injected with dehydroascorbic acid as well as those injected with ascorbic acid (Fig. 1B) ⇓ .
Fig. 1.
Vitamin C is transported into tumors independent of the redox state of the vitamin injected. A, mice were injected into the tail vein with [14C]ascorbic acid, [14C]dehydroascorbic acid, or [3H]sucrose and sacrificed 2 min after injection. Columns, means of 12 animals per group; bars, SE. B, HPLC analysis of the methanol-soluble fraction of the xenograft tumor (MDA468) of a mouse injected with 30 ��Ci of [14C]dehydroascorbic acid and sacrificed at 1 min. Accumulation of vitamin C in the tumor is in the form of ascorbic acid (AA; ∼91%; retention time, ∼7.93 min) in the dehydroascorbic acid (DHA)-injected animal
Vitamin C is taken up in the oxidized form as dehydroascorbic acid by most cells, including cancer cells, via the facilitative hexose transporters (GLUTs; Refs. 8, 9, 10 ). Thus, ascorbic acid must be oxidized in the pericellular milieu to dehydroascorbic acid to be transported through the GLUTs into the cell, where it is subsequently reduced and trapped in the form of ascorbic acid. Because most tumor cells are unable to transport ascorbic acid, we hypothesized that the injected ascorbic acid was oxidized to dehydroascorbic acid in the tumor microenvironment and then transported via the GLUTs. To ascertain whether ascorbic acid entered the tumor cells in this manner, we conducted inhibition experiments with d- and l-glucose because GLUTs selectively transport d-glucose but not l-glucose. d-Deoxyglucose and d-glucose have been shown to inhibit uptake of dehydroascorbic acid in the brain through GLUTs up to 70% in a dose-dependent fashion (11) . In the tumor-bearing mice, d-deoxyglucose inhibited the transport and accumulation of both dehydroascorbic acid and ascorbic acid (Fig. 2, A�CD) ⇓ . The 5-s time point was used to study transport, and there was an approximate maximal 35�C50% dose-dependent inhibition of vitamin C uptake (both injected dehydroascorbic acid and ascorbic acid). The 1- and 5-min time points represented both transport and accumulation and demonstrated an approximate dose-dependent 40�C65% maximal inhibition of vitamin C uptake (both injected dehydroascorbic acid and ascorbic acid). Administration of l-glucose had no effect on vitamin C transport or accumulation in the xenografts (data not shown). The inhibition by deoxyglucose of tumor vitamin C uptake and accumulation, in mice injected with ascorbic acid, indicated that the ascorbic acid was converted to dehydroascorbic acid and thereby transported into the tumor cells by the GLUTs.
��
Fig. 2.
Specificity of the transport of dehydroascorbic acid through GLUT1 in the tumor xenografts. [14C]Dehydroascorbic acid and [14C]ascorbic acid entered the xenograft tumors (A, LNCaP; B, HL-60; C, CEM; and D, MDA468), and their accumulation was blocked by increasing amounts of d-deoxyglucose, which is transported through GLUT1 (▪, 0 mg of d-deoxyglucose; , 5.3 mg of d-deoxyglucose; Embedded Image, 13.4 mg of d-deoxyglucose). A mouse has a baseline blood glucose concentration of ∼5�C7 mm or 2.67 mg of glucose in the entire animal, based on an average blood volume of the mouse. The amount of exogenous glucose administered in this experiment was based on whole mouse blood glucose and subsequent multiples to a maximum tolerable amount. The measured concentrations of both blood glucose and dehydroascorbic acid in the mice change rapidly and widely during the time after injection and it was difficult to obtain meaningful quantitative data regarding blood levels. Serum concentrations of ascorbic acid and dehydroascorbic acid were not affected by increasing concentrations of d-deoxyglucose or l-glucose. All experiments were carried out over the indicated time course. Columns, means of four mice; bars, SE.
��
We hypothesized that ascorbic acid was oxidized in the tumor microenvironment by superoxide anion. To test this concept, we coinjected animals bearing xenografts with ascorbic acid and SOD, catalase, or saline. The animals receiving SOD and radiolabeled ascorbic acid had an ∼50% reduction in tumor vitamin C accumulation, whereas there was no change in the tumor accumulation of vitamin C in animals coinjected with dehydroascorbic acid and SOD (Fig. 3A) ⇓ . There was no effect of coadministration of ascorbic acid and catalase, indicating that peroxide likely did not play a role in oxidizing ascorbic acid to dehydroascorbic acid (Fig. 3A) ⇓ .
Fig. 3.
Superoxide anions are responsible for the oxidation of vitamin C in the tumor xenografts. A, [14C]dehydroascorbic acid and [14C]ascorbic acid both entered the tumor xenografts and the accumulation of ascorbic acid, by the xenografts, was blocked by coinjected SOD and not by catalase or saline. All experiments were carried out over 1 min. Columns, means of four mice; bars, SE. B, superoxide anions generated by the tumor microenvironment are responsible for the oxidation of vitamin C in the tumor xenografts. All experiments were carried out over 60 min. Columns, means of four replicates; bars, SE.
We tested the ability of the tumor cells themselves to generate superoxide anion. Production of superoxide anion was only demonstrable with the HL-60 cell line (Fig. 3B) ⇓ , and the other cell lines had no capacity to generate superoxide anion. As expected, the HL-60 cell line increased superoxide generation when activated with PMA, whereas the other cell lines showed no PMA-inducible superoxide anion generation. Even the minced HL-60 xenograft tumors had a 3-fold increase in superoxide anion generation compared with the parent HL-60 cell line. Because minced xenograft tumors, distinct from the cell lines, had a prominent ability to generate superoxide anion, we concluded that nonneoplastic cells in the tumor stroma were responsible for the superoxide generation.
Discussion
We sought to determine how human tumor cells take up vitamin C in vivo by using human tumor xenograft models in nude athymic mice and measuring the uptake of [14C]ascorbic acid. The tumors rapidly took up the label after injection of ascorbic acid, although we had previously shown in vitro that the cells were unable to directly transport ascorbic acid (8 , 10) . We postulated that the tumor cells were taking up vitamin C in the form of dehydroascorbic acid. The observed inhibition of uptake by injected deoxyglucose confirmed the notion that the cells obtained vitamin C in the form of dehydroascorbic acid transported through the facilitative GLUTs. Because the GLUTs only transport the oxidized form of vitamin C and dehydroascorbic acid and because the cells had no other mechanism for taking up vitamin C, it was apparent that the ascorbic acid was oxidized to dehydroascorbic acid in the tumor microenvironment. The simultaneous lack of uptake of 14C label by the brain indicated that the ascorbic acid was not oxidized in the circulation. The inhibition of vitamin C uptake in ascorbic acid-injected animals by SOD pointed to a role for superoxide anion in the oxidation of ascorbic acid. The lack of production of superoxide anion by the epithelial tumors and the presence of superoxide anion generation in xenograft tumors suggested stromal cells as the source of the superoxide anion. We propose, therefore, that ascorbic acid is oxidized in the tumor microenvironment through the production of superoxide anion by nonmalignant stromal cells. Constitutive superoxide anion production has been demonstrated by fibroblasts, neutrophils, monocytes, and endothelial cells that have the NADPH oxidase enzyme (14, 15, 16, 17) . Neutrophils and HL-60 cells have been shown to oxidize extracellular ascorbic acid in a superoxide-dependent fashion (16 , 18 , 19) . Certain tumors (e.g., myeloid leukemia, the HL-60 xenograft in this study) can oxidize ascorbic acid directly, whereas others (epithelial tumors) may rely on nonneoplastic stromal cells for this function. A model for the transport and accumulation of vitamin C by tumors is summarized in Fig. 4 ⇓ .
Fig. 4.
A proposed mechanism of vitamin C transport and accumulation in tumors. Ascorbic acid, the predominant form of vitamin C in blood, is oxidized to dehydroascorbic acid in the tumor microenvironment by superoxide anion. Oxidation is, thus, a regulatory step for tumor transport of vitamin C. The dehydroascorbic acid is then transported through GLUT1 at the surface of the tumor cell. The dehydroascorbic acid in the tumor is then reduced to ascorbic acid, which is trapped in the tumor because it cannot be transported through GLUT1.
A sodium ascorbate cotransporter is present in many organs (12) although we have found no sodium-dependent ascorbic acid uptake in the neutrophils, monocytes, HL-60 cells, T-lymphocyte cell lines, or prostate and breast cancer cell lines included in this study (9 , 10) . Thus, the uptake of vitamin C in the form of dehydroascorbic acid through the GLUTs appears to be a general mechanism for vitamin C uptake. As we have shown previously, the sodium ascorbate cotransporter may have a role in vitamin C uptake of certain tumors (6) .
The precise function of vitamin C in neoplastic cells is not known. Studies have demonstrated increased concentrations of vitamin C in certain human tumors compared with normal tissue (7) , and vitamin C is important for the growth of some neoplastic cells in vitro (20) . Increased vitamin C consumption will raise the serum ascorbic acid level (21) and, therefore, could be expected to increase the tumor concentration of vitamin C. The increased intracellular concentration of vitamin C may have effects on tumor growth and the tumor��s ability to respond to oxidative stress associated with chemotherapy and radiation therapy. Although studies evaluating the role of vitamin C supplementation in cancer patients have generally shown no benefit with respect to survival or tumor regression (22) , it is not known whether high concentrations of vitamin C in human tumors afford the malignant cells with a metabolic advantage.
http://cancerres.aacrjournals.org/content/59/18/4555
.png)
.png)
.png)
.png)
.png)
