��
�����ڵĹ�������
Hydrogen peroxide in the human body
Abstract
Hydrogen peroxide (H2O2) is widely regarded as a cytotoxic agent whose levels must be minimized by the action of antioxidant defence enzymes. In fact, H2O2 is poorly reactive in the absence of transition metal ions. Exposure of certain human tissues to H2O2 may be greater than is commonly supposed: substantial amounts of H2O2 can be present in beverages commonly drunk (especially instant coffee), in freshly voided human urine, and in exhaled air. Levels of H2O2 in the human body may be controlled not only by catabolism but also by excretion, and H2O2 could play a role in the regulation of renal function and as an antibacterial agent in the urine. Urinary H2O2 levels are influenced by diet, but under certain conditions might be a valuable biomarker of ��oxidative stress��.
Previous article in issueNext article in issue
Keywords
Hydrogen peroxideHydroxyl radicalUrineBreathCatalaseOxygen electrode
1. Introduction
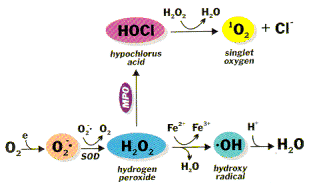
Hydrogen peroxide (H2O2) is a pale-blue covalent liquid, freely miscible with water and apparently able to cross cell membranes readily, although the pathways it uses to traverse have not been elucidated [1]. Multiple papers have described high (usually ��50 ��M) levels of H2O2 as being cytotoxic to a wide range of animal, plant and bacterial cells in culture, although LD50 values and the mode of cell death induced (apoptosis or necrosis) depend on the cell type used, its physiological state, length of exposure to H2O2, the H2O2 concentration used, and the cell culture media employed [1], [2], [3], [4], [5]. It is therefore widely thought that H2O2 is very toxic in vivo and must be rapidly eliminated, employing enzymes such as catalases, peroxidases (especially glutathione peroxidases) and thioredoxin-linked systems [1], [6], [7], [8], [9]. Paradoxically, however, acatalasemia in humans [1] appears to produce no significant phenotype, nor does ��knockout�� of glutathione peroxidase in mice except under certain conditions of abnormally high oxidative stress [10], [11], [12], [13].
In chemical terms, H2O2 is poorly reactive: it can act as a mild oxidizing or as a mild reducing agent, but it does not oxidize most biological molecules readily, including lipids, DNA and proteins (unless the latter have hyper-reactive thiol groups or methionine residues [1], [3], [14]). The danger of H2O2 largely comes from its ready conversion to the indiscriminately reactive hydroxyl radical (OH), either by exposure to ultraviolet light [15]
H2O2��uv2OH
or by interaction with a range of transition metal ions, of which the most important in vivo is probably iron [1], [16]
Fe2++H2O2��intermediate complexes (ferryl??)��Fe3++OH+OH−
Living organisms have evolved mechanisms to sequester transition metal ions into protein-bound forms that cannot catalyze OH formation and other free radical reactions in vivo. These mechanisms are especially important in such extracellular fluids as the blood plasma [1], [16], [17]. Nevertheless, H2O2 can contribute to Fenton chemistry not only by being one of the substrates but also by providing the other, e.g. by liberating iron from heme proteins [1], [16], [17], [18], [19]. Addition of H2O2 to cells in culture can lead to transition metal ion-dependent OH-mediated oxidative DNA damage, although this damage appears to be rapidly repaired provided that the cells are not rendered non-viable by an excess of H2O2[20].
However, levels of H2O2 at or below about 20�C50 ��M seem to have limited cytotoxicity to many cell types. Indeed, there is a growing literature showing that H2O2 can be used as an inter- and intra-cellular signalling molecule [21], [22], [23], [24], [25], [26]. The first example to be elucidated was the role of H2O2 as a second messenger in the activation of NF��B in some [23], but not all [27], cell types. Other examples of signalling roles for H2O2 have accumulated fast [21], [22], [23], [24], [25], [26]. Hence these may be a good reason not to eliminate all the H2O2 generated in vivo; its use in physiological signalling mechanisms. At sites of inflammation, H2O2 generated by activated phagocytes appears to modulate the inflammatory process, e.g. by up-regulating expression of adhesion molecules, controlling cell proliferation or apoptosis and modulating platelet aggregation [3], [4], [28], [29], [30], [31], [32], [33].
2. Exposure of human tissues to H2O2
Hydrogen peroxide is generated in vivo by the dismutation of superoxide radical (O2−), both non-enzymatically and catalyzed by superoxide dismutase enzymes. Hydrogen peroxide is also directly produced by a range of oxidase enzymes including glycollate and monoamine oxidases as well as by the peroxisomal pathway for ��-oxidation of fatty acids [1], [6], [34], [35]. Transgenic mice lacking mitochondrial superoxide dismutase suffer severe pulmonary and neurological damage, indicating the essentiality of removing intra-mitochondrial O2− in vivo [36], [37], [38]. However, with the apparent exception of cardiac muscle, mitochondria in most tissues appear to have limited capacity to remove H2O2, in that they readily generate substantial amounts of H2O2 in vitro and probably in vivo [1], [6], [35], [39], [40], [41]. Although mitochondria contain glutathione peroxidase and thioredoxin-linked peroxidase activities [42], [43], [44], the efficiency of these enzymes in removing H2O2 is uncertain given the ease with which mitochondria release H2O2[1], [6], [39], [40], [41].
It thus seems likely that most or all human cells are exposed to some level of H2O2, with the mitochondria being an important source. However, certain tissues may be exposed to higher H2O2 concentrations.
2.1. The oral cavity, oesophagus and stomach
Several beverages commonly drunk by humans can contain H2O2 at concentrations above 100 ��M, including green and black tea and especially instant coffee [45], [46], [47]. When such beverages are ingested, the H2O2 they contain presumably rapidly diffuses into the cells of the oral cavity and upper part of the gastrointestinal tract [48]. Oral bacteria also produce H2O2[49], [50], although the resulting levels of exposure of the oral tissues are uncertain. It is often suggested that H2O2 released into saliva is used by salivary peroxidase to oxidize thiocyanate (CNS−) into products toxic to certain bacterial strains [50].
2.2. The respiratory system
The cells lining the respiratory system, in common with the oral and oesophageal epithelium, are exposed to high O2 concentrations (21%) as compared with most other body tissues [1]. Hydrogen peroxide is present in exhaled air of humans [51], [52], [53], [54], [55], [56], [57], [58], [59] and of rats [60], although it is uncertain whether this H2O2 originates from oral bacteria [49], [50], phagocytes (e.g. alveolar macrophages, neutrophils in the oral cavity, or neutrophils recruited to the lungs in inflammatory lung diseases) or other lung cells. Amounts of exhaled H2O2 appear greater in subjects with inflammatory lung diseases [52], [53], [54], [55], [56], [57], [58] and in cigarette smokers [59]. Nevertheless, H2O2 is present in the air exhaled by healthy human subjects [51], [52], [53], [54], [55], [56], [57], [58], [59].
2.3. The kidney, urinary tract and bladder
Substantial quantities of H2O2, at concentrations sometimes exceeding 100 ��M, can be detected in freshly voided human urine (Table 1) [61], [62], [63], even in babies [64]. The simplest way of demonstrating its presence is to place urine into an oxygen electrode, and inject catalase through the cap. A ��spike�� of O2 release results as the H2O2 present is decomposed by catalase [63].
Table 1. Levels of hydrogen peroxide in freshly voided human urine
Gender of subject Age (years) [H2O2] in urine (��M)
Female 18 5.0
Female 19 8.0
Female 19 0.4
Female 21 6.2
Female 22 7.7
Female 25 11.5
Female 27 13.0
Female 35 3.5
Male 20 26.5
Male 21 16.3
Male 23 5.2
Male 26 5.9
Male 28 18.9
Male 30 22.3
Male 34 11.0
Male 49 109.6
Spot urine samples were collected from healthy human volunteers and assayed immediately. Subjects undertook no special dietary or other preparation before providing samples. Data are means of replicate determinations on each sample; replicates varied by <5%. H2O2 was analyzed by the ferrous ion oxidation�Cxylenol orange assay. Some data abstracted from [63], the rest provided by Caroline Manonmani, Mangala Srinivas, Melissa Sim and Yogeshwar Emritoll, students enrolled in the Talent Development Programme of the National University of Singapore.
The H2O2 detected in human urine appears to arise, at least in part, by O2−-dependent autoxidation of urinary molecules, some of which originate from diet [47], [63], [65]. Traces of superoxide dismutase are present in urine [66]: this enzyme, as well as the acidic pH of urine, should facilitate both enzymic and non-enzymic dismutation of O2− to H2O2[1]. The pO2 of urine within the bladder is below that of ambient air [67], [68] and so the rate of H2O2 generation in urine may well increase upon voiding. Nevertheless, the high levels of H2O2 that can be detected in some urine samples (Table 1) strongly suggest that at least some H2O2 generation occurs within the bladder. Indeed, H2O2 has been detected in urine sampled by catheterization [69]. Hydrogen peroxide has an antibacterial effect [1], [2], [70] and it may be that its presence at high levels in urine could be advantageous in diminishing infections of the bladder and urinary tract. On the other hand, the impact of H2O2 generation in vivo upon the cells lining the kidney tubules, ureters, bladder and urinary tract must be considered. Indeed, there are suggestions that H2O2 is involved in modulation of renal function [71], [72], [73]. Another possibility is that excretion of H2O2 represents a metabolic mechanism for controlling its levels in the human body. If so, measurement of urinary H2O2 levels may represent a valuable tool for assessment of ��oxidative stress��, since H2O2 can be measured rapidly and simply [63], [65]. This suggested route of H2O2 elimination by excretion is perhaps analogous to certain fish, which appear to dispose of H2O2 by excreting it through their gills [74].
2.4. Vascular endothelial and circulating blood cells
Some studies have claimed substantial levels of H2O2 (up to ∼35 ��M) in human blood plasma [75], [76], [77], but others have claimed levels to be very low, at or close to zero [78]. The latter data seem more credible, since H2O2 added to human plasma disappears rapidly. In part, it is degraded by the traces of catalase present, but H2O2 can also react with heme proteins, ascorbate, and protein-SH groups [1], [79]. In vivo, H2O2 generated in plasma could also diffuse into erythrocytes, white cells, endothelial cells and platelets for metabolism. However, the studies in [75], [76], [77] could be interpreted to suggest that H2O2 can be detected at high levels in plasma under assay conditions in which its removal is prevented. This implies that human plasma may be continuously generating H2O2. One enzyme involved in this process, at least under pathological conditions, appears to be xanthine oxidase [80]. Levels of circulating and endothelium-bound xanthine oxidase are increased as a result of tissue injury [81], [82].
2.5. Ocular tissues
The presence of H2O2, at widely varying levels (in some cases, 100 ��M or more), has been reported in human and other animal aqueous and vitreous humors [83], [84]. The explanation might be essentially the same as that advanced above to account for the conflicting data reported for blood plasma, i.e. that ocular fluids constantly generate H2O2, which is rapidly removed [83]. Any impairment in the capacity of the lens epithelium, retina or other ocular tissues to dispose of H2O2 would then result in its accumulation. The origin of this H2O2 is uncertain, but oxidation of glutathione or ascorbate is one possibility [84].
3. Conclusion
Hydrogen peroxide appears to be a ubiquitous molecule. We exhale it, excrete it and take it in from diet. It can be detected in drinking water, rain water and sea water [85], [86], [87], [88], [89]. These data emphasize the importance of metal ion sequestration in preventing the toxicity of H2O2 in vivo by decreasing the occurrence of Fenton chemistry, and help explain why a failure of such sequestration can produce devastating tissue damage in almost all organs of the body [1], [16].
FEBS Letters
Volume 486, Issue 1, 1 December 2000, Pages 10-13
��
Edited by Matti Saraste
Author links open overlay panelBarryHalliwellaMarie VeroniqueClementbLee HuaLonga
Show more
https://doi.org/10.1016/S0014-5793(00)02197-9Get rights and content
Under an Elsevier user licenseopen archive
��
Hydrogen peroxide in the human body - ScienceDirect https://www.sciencedirect.com/science/article/pii/S0014579300021979
https://www.mcgill.ca/oss/article/general-science-you-asked/hydrogen-peroxide-bodys-best-defence-system
��
��