��
������ ����֮�����ٲ�֮Դ The Gut is the base of Health and The Root of Disease
��
1. zinc
2. Omega-3,ALA, EPA, DHA
3. diarrhea��calcium + vitamin D
��
��
a Prominent health conditions with both biologic age and chronic inflammation as central risk factors. b Prominent health conditions with evidence linking them to gut dysbiosis. Note the similarities between the conditions associated with aging and inflammation and those associated with gut dysbiosis
Simplified schematic outlining the potential relationships among health risk factors, gut dysbiosis, inflammation, and age-related disease��as well as potential interventions for attenuating gut-associated inflammation
(Dis)Trust your gut: the gut microbiome in age-related inflammation, health, and disease | Microbiome | Full Text
https://microbiomejournal.biomedcentral.com/articles/10.1186/s40168-017-0296-0��
Control Secretory IgA & Keep A Happy Gut �� MyBioHack | Unlock Your Maximum Potential
https://mybiohack.com/blog/siga-secretory-iga-glutamine-tgf-il-6-10-microbiome-homeostasis-inflammation-high-low
��
Control Secretory IgA & Keep A Happy Gut �� MyBioHack | Unlock Your Maximum Potential
https://mybiohack.com/blog/siga-secretory-iga-glutamine-tgf-il-6-10-microbiome-homeostasis-inflammation-high-lowM Cells (Microfold Cells)
Microfold or M cells are epithelial cells embedded in the epithelium of mucosal tissues such as intestine, lung, and nasopharynx. Their name derives from the characteristic morphology of their apical or lumen-facing surface which has few if any microvilli.
The importance of M cells is that they are a critical player in the mucosal immune response and immune surveillance. The have have a unique ability to take up a variety of antigens - bacteria, viruses, parasites and proteins - from the lumen, transport them by transcytosis across the cell and deliver them to immune cells lying underneath, particularly dendritic cells. In other words, M cells are antigen-delivery cells for the immune system in places like the intestine and lung, the first step in eliciting a mucosal immune response. The capture of lumenal antigens occurs through electrostatic interactions and by binding to a number of adhesion molecules on the surface of the M cell.
M cells are most abundant in the epithelium overlying lymphoid follicles such as Peyer's patches in the intestine. There are additional types of M cells such as villous M cells interspersed among entercytes over villi in the small intestine.
The ability of M cells to deliver antigens to the mucosal immune system has led to the concept of exploiting the cells for delivery of vaccines targeting mucosal immune responses, which is an active area of research.
Paradoxically, although M cells are important in generating an effective mucosal immune response to protect against many pathogens, they also serve as an entry portal for a number of pathogens that can be present in the intestinal lumen. Examples include bacteria such as Brucella, Yersinia, and Salmonella species, viruses like reoviruses and poliovirus, and prions. In these cases, the pathogen is taken up and sometimes replicates in M cells, and is then transferred across the cell to induce a systemic infection.M Cells
http://www.vivo.colostate.edu/hbooks/pathphys/digestion/smallgut/mcells.html��
Relationship between microbiota diversity, epithelial Integrity, and inflammation in disease state
��

��
The lysis of gram-negative bacteria causes them to release lipopolysaccharide (LPS; endotoxin) from the outer membrane of their cell wall. The LPS binds to a LPS-binding protein circulating in the blood and this complex, in turn, binds to a receptor molecule (CD14) found on the surface of body defense cells called macrophages. This is thought to promote the ability of the toll-like receptor TLR-4 to respond to the LPS, triggering the macrophage to release various defense regulatory chemicals called cytokines, including IL-1, IL-6, IL-8, TNF-alpha, and PAF. The cytokines then bind to cytokine receptors on target cells and initiate inflammation and activate both the complement pathways and the coagulation pathway. (LPS, lipopolysaccharide; IL-1, interleukin-1; IL-6, interleukin-6; IL-8, interleukin-8, TNF-alpha, tumor necrosis factor-alpha; PAF, platelet-activating factor.) This will be discussed in greater detail under Bacterial Pathogenicity.
Another cell surface PRR is CD14. CD14 is found on monocytes, macrophages, and neutrophils and promotes the ability of TLR-4 to respond to LPS. LPS typically binds to LPS-binding protein in the plasma and tissue fluid. The LPS-binding protein promotes the binding of LPS to the CD14 receptors. At that point the LPS-binding protein comes off and the LPS-CD14 bind to TLR-4. Interaction of LPS and CD14 with TLR-4 leads to an elevated synthesis and secretion of inflammatory cytokines such as IL-1, IL-6, IL-8, TNF-alpha, and platelet-activating factor (PAF). These cytokines then bind to cytokine receptors on target cells and initiate inflammation and activate both the complement pathways and the coagulation pathway (see Fig. 7).Innate Immunity
Early Induced Innate Immunity: Pattern-Recognition Receptors (PRRs) and Danger-Recognition Receptors (DRRs)
Fundamental Statement for this Softchalk Lesson:
1. Early induced innate immunity begins 4 - 96 hours after exposure to an infectious agent and involves the recruitment of defense cells as a result of pathogen-associated molecular patterns or PAMPS binding to pattern-recognition receptors or PRRs and danger-associated molecular patterns or DAMPs binding to danger-recognition receptors or DRRs.
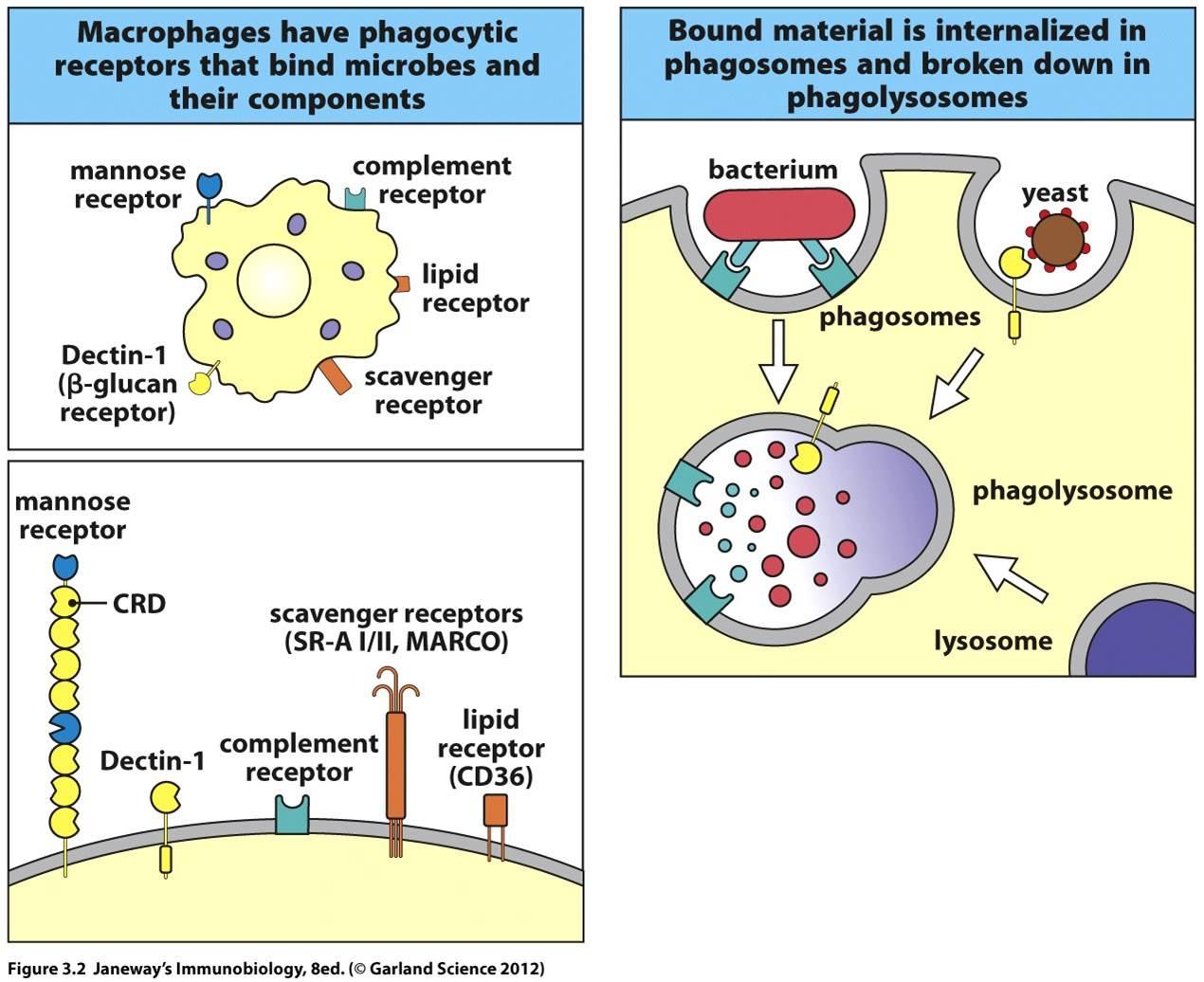
2. Endocytic pattern-recognition receptors are found on the surface of phagocytes and promote the attachment of microorganisms to phagocytes leading to their subsequent engulfment and destruction. They include mannose receptors, scavenger receptors, and opsonin receptors.
3. Binding of microbial PAMPs to signaling PRRs promotes the production of inflammatory cytokines, antiviral cytokines called type-1 interferons (IFN), chemotactic factors, and antimicrobial peptides. They include toll-like receptors (TLRs) and NODs.
4. PRRs found on the surface of the body's cells typically bind to surface PAMPs on microbes and stimulate the production of inflammatory cytokines.
5. PRRs found within cellular phagolysosomes (endosomes) typically detect nucleic acid PAMPs released during the phagocytic destruction of viruses and stimulate the production of antiviral cytokines called type-1 interferons.
6. PRRs and DRRs found within the cytoplasm of host cells typically trigger the formation of multi-protein complexes called inflammasomes which, in turn, triggers the formation of inflammatory cytokines and can also leads to an inflammatory response-induced cell suicide called pyroptosis.
7. PRRs circulating in the blood and tissue fluid activate the complement pathways and may function as opsonins.
��
�����յ����������ߣ�ģʽʶ�����壨PRR����Σ��ʶ�����壨DRR����1.�����յ����������߿�ʼ�ڱ�¶�ڴ�Ⱦ����4-96Сʱ�����ڲ�ԭ����ط���ģʽ��PAMPS��ģʽʶ�������PRR����Լ�Σ����ط���ģʽ��DAMP��Σ��ʶ�������DRR��ϡ�
2.������ϸ���ı��淢������ģʽʶ�����壬���ٽ�����������ϸ���ĸ��ţ�������������ɺ��ƻ������ǰ�����¶�����壬���������͵�����(opsonins)���塣
3.����PAMP���ź�PRR�Ľ�ϻ�ٽ���֢ϸ�����ӣ���Ϊ1�����أ�IFN���Ŀ�����ϸ�����ӣ��������ӺͿ����ĵIJ��������ǰ����շ������壨TLR����NOD��
4.������ϸ�����淢�ֵ�PRRͨ������������PAMP��ϲ��̼�����ϸ�����ӵIJ�����
5.��ϸ��������ø�壨���壩�з��ֵ�PRRͨ����ⲡ�������ƻ��������ͷŵĺ���PAMP�����̼���Ϊ1�����صĿ�����ϸ�����ӵIJ�����
6.������ϸ�����з��ֵ�PRR��DRRͨ���ᴥ����Ϊ��֢С��Ķ൰��������γɣ�����������֢��ϸ�����ӵ��γɣ������ܵ�����֢��Ӧ�յ���ϸ����ɱ����Ϊ�����ữ��
7.��ѪҺ����֯Һ��ѭ����PRR�����;����������������ص����á�
��Early Induced Innate Immunity: Pattern-Recognition Receptors (PRRs) and Danger-Recognition Receptors (DRRs)
Early induced innate immunity begins 4 - 96 hours after exposure to an infectious agent and involves the recruitment of defense cells as a result of pathogen-associated molecular patterns or PAMPS binding to pattern-recognition receptors or PRRs. These recruited defense cells include:
phagocytic cells: leukocytes such as neutrophils, eosinophils, and monocytes; tissue phagocytic cells in the tissue such as macrophages;
cells that release inflammatory mediators: inflammatory cells in the tissue such as macrophages and mast cells; leukocytes such as basophils and eosinophils; and
natural killer cells (NK cells).
Unlike adaptive immunity, innate immunity does not recognize every possible antigen. Instead, it is designed to recognize molecules shared by groups of related microbes that are essential for the survival of those organisms and are not found associated with mammalian cells. These unique microbial molecules are called pathogen-associated molecular patterns or PAMPS and include LPS from the gram-negative cell wall, peptidoglycan and lipotechoic acids from the gram-positive cell wall, the sugar mannose (a terminal sugar common in microbial glycolipids and glycoproteins but rare in those of humans), bacterial and viral unmethylated CpG DNA, bacterial flagellin, the amino acid N-formylmethionine found in bacterial proteins, double-stranded and single-stranded RNA from viruses, and glucans from fungal cell walls. In addition, unique molecules displayed on stressed, injured, infected, or transformed human cells also be recognized as a part of innate immunity. These are often referred to as danger-associated molecular patterns or DAMPs.
Most body defense cells have pattern-recognition receptors or PRRs for these common PAMPS (see Fig. 1) and so there is an immediate response against the invading microorganism. Pathogen-associated molecular patterns can also be recognized by a series of soluble pattern-recognition receptors in the blood that function as opsonins and initiate the complement pathways. In all, the innate immune system is thought to recognize approximately 103 of these microbial molecular patterns.�����յ����������ߣ�ģʽʶ�����壨PRR����Σ��ʶ�����壨DRR��
�����յ������������ڽӴ���Ⱦ����4-96Сʱ��ʼ�����ڲ�ԭ����صķ���ģʽ��PAMPS��ģʽʶ�������PRR��ϣ���ļ�˷���ϸ������Щ��ļ�ķ�����λ������
����ϸ������ϸ����������������ϸ������������ϸ���͵���ϸ������֯�е�����ϸ����֯���������ϸ����
�ͷ����Խ��ʵ�ϸ������֯�е�����ϸ�����������ϸ���ͷʴ�ϸ������ϸ�������ȼ�����ϸ������������ϸ������
��Ȼɱ��ϸ����NKϸ������
����Ӧ�����߲�ͬ���������߲���ʶ�����п��ܵĿ�ԭ��ȡ����֮���ǣ��������Ϊʶ�����������Ⱥ�����ķ��ӣ���Щ���Ӷ�����Щ�����������������Ҫ������δ�����벸�鶯��ϸ����صķ��ӡ���Щ���ص�������ӱ���Ϊ��ԭ����ط���ģʽ��PAMPS���������Ը���������ϸ���ڵ�LPS�����Ը���������ϸ���ڵ��ľ��Ǻ�֬���ᣬ�Ǹ�¶�ǣ�������֬���ǵ����г�����ĩ���ǣ���ϸ���Ͳ���δ������CpG DNA��ϸ����ë���ף�ϸ���������з��ֵİ�����N-���������ᣬ������˫���͵���RNA�Լ����ϸ���ڵ��Ͼ��ǵȡ����⣬��Ӧ�������ˣ���Ⱦ��ת��������ϸ����չʾ�Ķ��ط���Ҳ����Ϊ���������ߵ�һ���֡���Щͨ����Ϊ��Σ����صķ���ģʽ��DAMP��
������������ϸ��������Щ����PAMPS��ģʽʶ�������PRR����ͼ1������˿��Զ����ֵ���������������Ӧ����ԭ��صķ���ģʽҲ���Ա�ѪҺ�е�һϵ�п�����ģʽʶ��������ʶ����Щ����������ص����ò���������;������֮����������ϵͳ����Ϊ����ʶ���Լ103���������ģʽ��
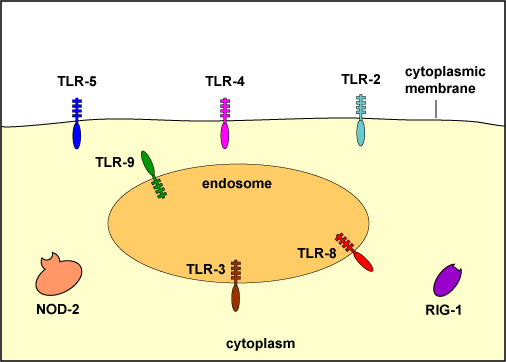
Glycoprotein molecules known as pattern-recognition receptors are found on the surface of a variety of body defense cells. They are so named because they recognize and bind to pathogen-associated molecular patterns - molecular components associated with microorganisms but not found as a part of eukaryotic cells. These include bacterial molecules such as peptidoglycan, teichoic acids, lipopolysaccharide, mannans, flagellin, pilin, and bacterial DNA. There are also pattern-recognition molecules for viral double-stranded RNA (dsRNA) and fungal cell walls components such as lipoteichoic acids, glycolipids, mannans, and zymosan. Many of these pattern recognition receptors are known as toll-like receptors.
�ڸ����������ϸ���ı��淢���˱���Ϊģʽʶ��������ǵ����ӡ�֮�����������ǣ�����Ϊ����ʶ����벡ԭ����صķ���ģʽ-��������صķ��ӳɷ֣�����δ��Ϊ���ϸ����һ���ֱ����֡���Щ����ϸ�����ӣ������ľ��ǣ������ᣬ֬���ǣ���¶���ǣ���ë���ף���ë����ϸ��DNA��Ҳ���ڲ���˫��RNA��dsRNA�������ϸ���ڳɷ֣�����֬���ף���֬����¶���Ǻͽ�ĸ���ǣ���ģʽʶ����ӡ���Щģʽʶ�������������౻��Ϊ�շ������塣
To recognize PAMPs such as those listed above, various body cells have a variety of corresponding receptors called pattern-recognition receptors or PRRs capable of binding specifically to conserved portions of these molecules. Cells that typically have pattern recognition receptors include macrophages, dendritic cells, endothelial cells, mucosal epithelial cells, and lymphocytes. Many pattern-recognition receptors are located on the surface of these cells where they can interact with PAMPs on the surface of microbes. Others PRRs are found within the phagolysosomes of phagocytes where they can interact with PAMPs located within microbes that have been phagocytosed. Some PRRs are found in the cytosol of the cell.
KEY
on the surface:
TLR-2 - recognizes peptidoglycan, bacterial lipoproteins, lipoteichoic acid, and porins
TLR-4 - recognizes lipopolysaccharide (LPS) from gram-negative cell wall, fungal mannans, viral envelope proteins, parasitic phospholipids, heat-shock proteins
TLR-5 - recognizes bacterial flagellin
within endosomes (phagolysosomes):
TLR-3 - recognizes viral double-stranded DNA
TLR-8 - recognizes viral single-stranded RNA
TLR-9 - recognizes viral and bacterial unmethylated CpG sequences
in the cytoplasm:
NOD-2 - recognizes muramyl dipeptide from bacterial peptidoglycan
RIG-1 - recognizes viral RNAMany pattern-recognition receptors are located on the surface of these cells where they can interact with PAMPs on the surface of microbes. Others PRRs are found within the phagolysosomes of phagocytes where they can interact with PAMPs located within microbes that have been phagocytosed. Some PRRs are found in the cytosol of the cell.
There are two functionally different major classes of pattern-recognition receptors: endocytic pattern-recognition receptors and signaling pattern-recognition receptors.��
a. Endocytic (Phagocytic) Pattern-Recognition Receptors
Endocytic pattern-recognition receptors, also called phagocytic pattern-recognition receptors, are found on the surface of phagocytes and promote the attachment of microorganisms to phagocytes leading to their subsequent engulfment and destruction. They include:
1. Mannose receptors
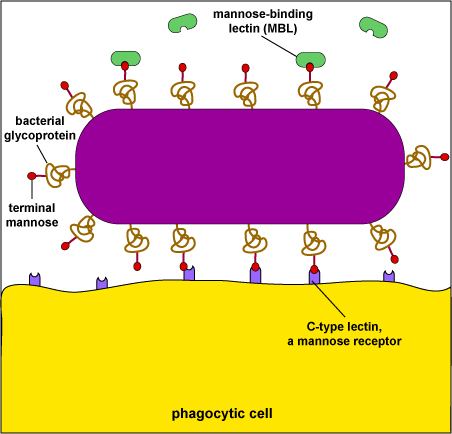
Mannose receptors on the surface of phagocytes bind to various microbial carbohydrates such as those rich in mannose or fucose, and to N-acetylglucosamine (NAG). Human glycoproteins and glycolipids typically have terminal N-acetylglucosamine and sialic acid groups. C-type lectins found on the surface of phagocytes are mannose receptors (see Fig. 3). It is now thought that mannose receptors may be quite important in removing potentially harmful mannose-containing glycoproteins such as lysosomal hydrolases that are produced in increased amounts during inflammation.
Mannose-rich glycans are short carbohydrate chains with the sugar mannose or fructose as the terminal sugar. They are commonly found in microbial glycoproteins and glycolipids but are rare in those of humans. (Human glycoproteins and glycolipids typically have terminal N-acetylglucosamine and sialic acid groups.) C-type lectins, found on the surface of phagocytes, are endocytic pattern recognition receptors that bind to mannose-rich glycans in order to attach microbes to phagocytes. Mannose-binding lectin (MBL), also known as mannan-binding protein, is a soluble pattern recognition receptor in plasma and tissue fluid that binds to mannose-rich glycans on microbes in oder to activate the lectin complement pathway.��
2. Dectin-1
Dectin-1 recognizes beta-glucans (polymers of gucose) commonly found in fungal cell walls.
3. Scavenger receptors
Scavenger receptors found on the surface of phagocytic cells bind to bacterial cell wall components such as LPS, peptidoglycan and teichoic acids (see Fig. 1). There are also scavenger receptors for certain components of other types of microorganisms, as well as for stressed, infected, or injured cells . Scavenger receptors include CD-36, CD-68, and SRB-1.
4. Opsonin receptors
Opsonins are soluble molecules produced as a part of the body's immune defenses that bind microbes to phagocytes. One portion of the opsonin binds to a PAMP on the microbial surface and another portion binds to a specific receptor on the phagocytic cell.
Acute phase proteins circulating in the plasma, such as:
mannose-binding lectin (also called mannose-binding protein) that binds to various microbial carbohydrates such as those rich in mannose or fucose, and to N-acetylglucosamine (NAG); and
C-reactive protein (CRP) that binds to phosphorylcholine portion of teichoic acids and lipopolysaccharides of bacterial and fungal cell walls. It also binds to the phosphocholine found on the surface of damaged or dead human cells.
Complement pathway proteins, such as C3b (see Fig. 4) and C4b recognize a variety of PAMPS.
Surfactant proteins in the alveoli of the lungs, such as SP-A and SP-D are opsonins.
During adaptive immunity, the antibody molecule IgG can function as an opsonin (see Fig. 5).Fig. 4: Enhanced Attachment of Bacteria to Phagocytes by the Opsonin C3b
When body defense pathways known as the complement pathways are activated, one of the beneficial defense proteins made is called C3b. One portion of C3b binds to bacterial surface proteins and another portion binds to C3b receptors such as CD-1 on phagocytes. The process of enhanced attachment is called opsonization.��
Fig. 5: Enhanced Attachment (Opsonization) of a Bacterium By Way Of the Antibody IgG
One of the functions of certain antibody molecules known as IgG is to stick antigens such as bacterial proteins and polysaccharides to phagocytes. The tips of the antibody, the Fab portion, have a shape that fits epitopes, portions of an antigen with a complementary shape. The stalkof the antibody is called the Fc portion and is able to bind to Fc receptors on phagocytes. Also, when body defense pathways known as the complement pathways are activated, one of the beneficial defense proteins made is called C3b. C3b binds by one end to bacterial surface proteins and by the other end to C3b receptors on phagocytes. The IgG and C3b are also known as opsonins and the process of enhanced attachment is also called opsonization.��
��
Fig. 7: Physiologic Action of Lipopolysaccharide (LPS) from the Gram-Negative Cell Wall
��
The lysis of gram-negative bacteria causes them to release lipopolysaccharide (LPS; endotoxin) from the outer membrane of their cell wall. The LPS binds to a LPS-binding protein circulating in the blood and this complex, in turn, binds to a receptor molecule (CD14) found on the surface of body defense cells called macrophages. This is thought to promote the ability of the toll-like receptor TLR-4 to respond to the LPS, triggering the macrophage to release various defense regulatory chemicals called cytokines, including IL-1, IL-6, IL-8, TNF-alpha, and PAF. The cytokines then bind to cytokine receptors on target cells and initiate inflammation and activate both the complement pathways and the coagulation pathway. (LPS, lipopolysaccharide; IL-1, interleukin-1; IL-6, interleukin-6; IL-8, interleukin-8, TNF-alpha, tumor necrosis factor-alpha; PAF, platelet-activating factor.) This will be discussed in greater detail under Bacterial Pathogenicity.
Another cell surface PRR is CD14. CD14 is found on monocytes, macrophages, and neutrophils and promotes the ability of TLR-4 to respond to LPS. LPS typically binds to LPS-binding protein in the plasma and tissue fluid. The LPS-binding protein promotes the binding of LPS to the CD14 receptors. At that point the LPS-binding protein comes off and the LPS-CD14 bind to TLR-4. Interaction of LPS and CD14 with TLR-4 leads to an elevated synthesis and secretion of inflammatory cytokines such as IL-1, IL-6, IL-8, TNF-alpha, and platelet-activating factor (PAF). These cytokines then bind to cytokine receptors on target cells and initiate inflammation and activate both the complement pathways and the coagulation pathway (see Fig. 7).��
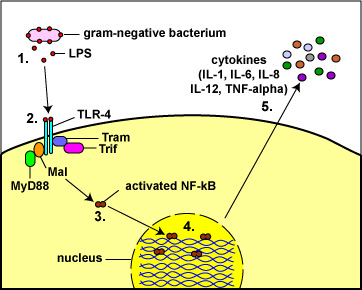
Fig. 8: Toll-Like Receptors Responding to Lipopolysaccharide (LPS)
from the Gram-Negative Cell Wall
(1) The lysis of Gram-negative bacteria causes them to release lipopolysaccharide (LPS; endotoxin) from the outer membrane of their cell wall.
(2) The LPS binds to a pair of TLR-4s on the macrophage.
(3&4) This enables regulatory molecules within the cell - Mal, MyD88, Tram, and Trif - to trigger reactions that activate a master regulator of inflammation called NF-kappa B. Activated NF-kappa B enters the cell's nucleus and switches on genes coding for cytokines such as:
a. Interleukin-1 (IL-1) and Tumor necrosis factor-alpha (TNF-alpha): enhance inflammatory responses;
b. Interleukin-8 (IL-8): aids in the ability of white blood cells to leave the blood vessels and enter the tissue; a chemoattractant for phagocytes;
c. Interleukin-6 (IL-6) promotes B-lymphocyte activity; and
d. Interleukin-12 (IL-12): promotes T-lymphocyte activity. (5)http://faculty.ccbcmd.edu/~gkaiser/SoftChalk%20BIOL%20230/Innate%20Immunity/PRRs/PRRs_print.html
Leaky Gut and Atopic Dermatitis: What Is the Connection?Tracing the skin as it rounds the lips and becomes the lining of the mouth and digestive tract, it seems reasonable that skin problems could continue on the inside as well. In fact, similar to the function of the skin, the lining of the intestines is tasked with keeping some things out (such as certain types of bacteria and larger molecules that may cause allergies) and allowing other things in (such as nutrients). An impairment of this function leads to increased permeability, sometimes called ��leaky gut."
Certain triggers and stresses are thought to cause changes in the intestinal bacteria and disrupt the gut barrier, and this has been tied to many different diseases including inflammatory bowel disease, celiac disease, food intolerance, and small intestinal bacterial overgrowth. Interestingly, there are also non-gastrointestinal diseases on this list, including fibromyalgia, chronic fatigue syndrome, and, the focus of the current article, atopic dermatitis.����Ƥ����ʹ��Χ���촽����Ϊ��ǻ���������ij�����Ƥ������Ҳ�������ڲ����������Ǻ����ġ�ʵ���ϣ�������Ƥ���Ĺ��ܣ����ڵ���������ֹijЩ�������루����ijЩ���͵�ϸ���Ϳ�����������Ĵ���ӣ��������������������루����Ӫ�������ù��ܵ�����ͨ�����ӣ���ʱ��Ϊ����©�Գ�����
������ΪijЩ�������غ�ѹ��������ϸ���ı仯���ƻ��������ϣ����������ͬ�ļ����йأ�������֢�Գ�������ǻ������ʳ�ﲻ���ܺ�С��ϸ��������������Ȥ���ǣ����б��л�������θ����������������ά��ʹ������ƣ���ۺ�֢���Լ����ĵ��ص�����Ӧ��Ƥ�ס���
Abnormal Barrier in Atopic Dermatitis and Leaky Gut
In atopic dermatitis, the skin barrier is known to be abnormal: water escapes too easily leading to dry, itchy skin, and allergens, irritants, and bacteria can all enter, leading to an immune response and subsequent inflammation. Although there is some evidence to show that excluding foods from the diet in those without known allergy is not helpful for most, the implication of the gut in atopic dermatitis has been very difficult to shake, and many feel that diet is the ��root cause.��
A study from the United Kingdom identified significantly increased intestinal permeability (leaky gut) in children with atopic dermatitis when compared to children without eczema, while another found a relationship between how leaky the gut was with how severe the eczema was--a very compelling finding, to be sure!
If leaky gut is indeed connected with atopic dermatitis, can it be treated? The answer seems to be ��yes,�� but with qualifications.��Ӧ��Ƥ�ͳ�©���쳣����
����Ӧ��Ƥ���У���֪Ƥ�������쳣��ˮ�������ݳ�������Ƥ��������������ҹ���ԭ���̼����ϸ�������Խ��룬�������߷�Ӧ��������֢��������֤�ݱ���������Щû�й���֢���˴���ʳ���ų�ʳ��Դ�����������£�����θ����Ӧ��Ƥ��Ӱ��ȴ���Ѷ�ҡ����������Ϊ��ʳ�ǡ�����ԭ�� ��
Ӣ����һ���о����֣���û��ʪ��Ķ�ͯ��ȣ���Ӧ��Ƥ�����ij���ͨ�ԣ�������©���������ӣ�����һ���о����֣�������©��ʪ������س̶�֮����ڹ���-����һ���dz������ŷ��ķ��֣� Ϊ��ȷ����
���������©ȷʵ����Ӧ��Ƥ���йأ����������𣿴��ƺ��ǡ��ǡ���������������
Neurotherapeutics
January 2018, Volume 15, Issue 1, pp 75�C91 | Cite as
Diet, Gut Microbiota, and Vitamins D + A in Multiple Sclerosis
1.Department of Sciences University of Basilicata Potenza Italy
Abstract
Central to the understanding of the relationships between diet, gut microbiota, and vitamins D and A in multiple sclerosis is low-grade inflammation, which is involved in all chronic inflammatory diseases and is influenced by each of the above effectors. We show that food components have either proinflammatory or anti-inflammatory effects and influence both the human metabolism (the ��metabolome��) and the composition of gut microbiota. Hypercaloric, high-animal-fat Western diets favor anabolism and change gut microbiota composition towards dysbiosis. Subsequent intestinal inflammation leads to leakage of the gut barrier, disruption of the blood�Cbrain barrier, and neuroinflammation. Conversely, a vegetarian diet, rich in fiber, is coherent with gut eubiosis and a healthy condition. Vitamin D levels, mainly insufficient in a persistent low-grade inflammatory status, can be restored to optimal values only by administration of high amounts of cholecalciferol. At its optimal values (>30 ng/ml), vitamin D requires vitamin A for the binding to the vitamin D receptor and exert its anti-inflammatory action. Both vitamins must be supplied to the subjects lacking vitamin D. We conclude that nutrients, including the nondigestible dietary fibers, have a leading role in tackling the low-grade inflammation associated with chronic inflammatory diseases. Their action is mediated by gut microbiota and any microbial change induced by diet modifies host�Cmicrobe interactions in a consequent way, to improve the disease or worsen it.
Keywords
Diet Gut Microbiota Vitamin D Multiple Sclerosis NeuroinflammationDiet, Gut Microbiota, and Vitamins D + A in Multiple Sclerosis | SpringerLink
https://link.springer.com/article/10.1007%2Fs13311-017-0581-4��
Published: 03 May 2019
The combination of sport and sport-specific diet is associated with characteristics of gut microbiota: an observational study
Lae-Guen Jang, Geunhoon Choi, Sung-Woo Kim, Byung-Yong Kim, Sunghee Lee & Hyon Park
Journal of the International Society of Sports Nutrition volume 16, Article number: 21 (2019)
Abstract
Background
Recently, gut microbiota have been studied extensively for health promotion, disease prevention, disease treatment, and exercise performance. It is recommended that athletes avoid dietary fiber and resistant starch to promote gastric emptying and reduce gastrointestinal distress during exercise, but this diet may reduce microbial diversity and compromise the health of the athlete��s gut microbiota.
Objective
This study compared fecal microbiota characteristics using high-throughput sequencing among healthy sedentary men (as controls), bodybuilders, and distance runners, as well as the relationships between microbiota characteristics, body composition, and nutritional status.
Methods
Body composition was measured using DXA, and physical activity level was assessed using IPAQ. Dietary intake was analyzed with the computerized nutritional evaluation program. The DNA of fecal samples was extracted and it was sequenced for the analysis of gut microbial diversity through bioinformatics cloud platform.
Results
We showed that exercise type was associated with athlete diet patterns (bodybuilders: high protein, high fat, low carbohydrate, and low dietary fiber diet; distance runners: low carbohydrate and low dietary fiber diet). However, athlete type did not differ in regard to gut microbiota alpha and beta diversity. Athlete type was significantly associated with the relative abundance of gut microbiota at the genus and species level: Faecalibacterium, Sutterella, Clostridium, Haemophilus, and Eisenbergiella were the highest (p < 0.05) in bodybuilders, while Bifidobacterium and Parasutterella were the lowest (p < 0.05). At the species level, intestinal beneficial bacteria widely used as probiotics (Bifidobacterium adolescentis group, Bifidobacterium longum group, Lactobacillus sakei group) and those producing short chain fatty acids (Blautia wexlerae, Eubacterium hallii) were the lowest in bodybuilders and the highest in controls. In addition, aerobic or resistance exercise training with an unbalanced intake of macronutrients and low intake of dietary fiber led to similar diversity of gut microbiota. Specifically, daily protein intake was negatively correlated with operation taxonomic unit (r = − 0.53, p < 0.05), ACE (r = − 0.51, p < 0.05), and Shannon index (r = − 0.64, p < 0.01) in distance runners..
Conclusion
Results suggest that high-protein diets may have a negative impact on gut microbiota diversity for athletes, while athletes in resistance sports that carry out the high protein low carbohydrates diet demonstrate a decrease in short chain fatty acid-producing commensal bacteria.The combination of sport and sport-specific diet is associated with characteristics of gut microbiota: an observational study | Journal of the International Society of Sports Nutrition | Full Text
https://jissn.biomedcentral.com/articles/10.1186/s12970-019-0290-y��
J Cell Biochem. 2018 Dec;119(12):9997-10004. doi: 10.1002/jcb.27329. Epub 2018 Aug 26.
LPS promotes the expression of PD-L1 in gastric cancer cells through NF-��B activation.
Li H1, Xia JQ1, Zhu FS1, Xi ZH1, Pan CY1, Gu LM1, Tian YZ1.
Author information
1
Department of Gastroenterology, Affiliated Hospital of Integrated Traditional Chinese and Western Medicine, Nanjing University of Chinese Medicine, Nanjing, Jiangsu, China.
Abstract
Gastric cancers are a group of highly aggressive malignancies with a huge disease burden worldwide. Gastric infections, such as helicobacter pylori, can induce the occurrence of gastric cancers. However, the role of gastric infection in gastric cancer development is unclear. Programmed death-ligand 1 (PD-L1, B7-H1) is a member of the B7 family of cell surface ligands, which binds the PD-1 transmembrane receptor and inhibits T-cell activation within cancer tissues. It has been reported that the expression of PD-L1 is inversely related to the prognosis of patients with gastric cancers. Therefore, the regulation of PD-L1 expression in gastric cancers needs to be studied. In the current study, we explored the possible effects of lipopolysaccharide (LPS) on PD-L1 expression in gastric cancer cells. We observed that LPS stimulation could markedly increase PD-L1 expression in gastric cancer cells. Furthermore, we found that nuclear factor-��B (NF-��B) activation was involved in PD-L1 expression in gastric cancer cells exposed to LPS stimulation through p65-binding to the PD-L1 promoter. Taken together, these data indicate that gastric infection might promote the development of gastric cancers thought the LPS-NF-��B-PD-L1 axis.
����
Jϸ�����ﻯѧ��2018��12��;119(12):9997-10004��doi: 10.1002 / jcb.27329��Epub 2018 8��26�ա�
LPS�ٽ�PD-L1�ı�����θ��ϸ��ͨ��NF-��B����
Li H1, Xia JQ1, Zhu FS1, Xi ZH1, Pan CY1, Gu LM1, Tian YZ1��
������Ϣ
1 �Ͼ���ҽҩ��ѧ����ҽ���ҽԺ�����ڿƣ������Ͼ���
ժҪ
θ����һ����Գ̶ȼ��ߵĶ��������������緶Χ�ھ��о�ļ���������θ��Ⱦ���������ݸ˾���Ⱦ��������θ���ķ�����Ȼ����θ��Ⱦ��θ��������չ�е������в���ȷ����������������1 (PD-L1, B7- h1)��ϸ����������B7����ij�Ա֮һ������PD-1��Ĥ�����ϣ����ư�ϸ����֯��Tϸ���Ļ���б�����PD-L1�ı�����θ�����ߵ�Ԥ��ʸ���ء���ˣ�PD-L1��θ���еı��������Ҫ��һ���о����ڱ��о��У�����̽����֬����(LPS)��θ��ϸ����PD-L1����Ŀ���Ӱ�졣���ǹ۲쵽LPS�̼�������������θ��ϸ����PD-L1�ı������,���Ƿ��ֺ�factor-��B (NF-��B)�������PD-L1������θ��ϸ����¶��LPS�̼�ͨ��p65-binding PD-L1�����ӡ���������,��Щ���ݱ���θ��Ⱦ���ܴٽ�θ��֢�ķ�չ��ΪLPS-NF-��B-PD-L1�ᡣ
©2018�����ڿ�����˾
© 2018 Wiley Periodicals, Inc.LPS promotes the expression of PD-L1 in gastric cancer cells through NF-��B activation. - PubMed - NCBI
https://www.ncbi.nlm.nih.gov/pubmed/30145830��
Dietary Emulsifier�CInduced Low-Grade Inflammation Promotes Colon Carcinogenesis
��ʳ�黯���յ��������֢�ٽ��᳦������
������ά��Ŭ�ߣ�Emilie Viennois�����ϵϰ���÷�֣�Didier Merlin��������³��T����ά�ȣ�Andrew T.
DOI��10.1158 / 0008-5472.CAN-16-1359������2017��1��
����ͼ����������Ϣ�����PDF
����
��֢�Գ�����IBD�������ֱ������չ�ķ������ӣ�����������᳦����ذ�������֢�ٽ��᳦���������ĸ����IBD�������ļ����ǵͶ���֢����������������ж����ڵij�������Ⱥ��ɸı�ʹ�л�ۺ����йء�����ķ��ֱ�����ʳ���黯���ɴٽ������Ͷ���֢����ʳ���黯���Ǽӹ�ʳƷ���ձ���ڵijɷ֣��ɸı䳦����Ⱥ����ɡ�����������ڽ᳦���շ��Ĵ����ٴ�ǰģ����֤��������ʳ����ʳ�黯�����ȼ���ά�ػ��ɽ����80��Ӿ������ķ�չ����ǿ��������չ��ı��������Ԫ�������йأ�����������֬���Ǻͱ�ë����ˮƽ���ߡ����Ƿ��֣��黯����������������ĸı��DZ�Ҫ�ģ���������������Ҫ����ֳ�͵����ź�ͨ·�ĸı䣬��Щͨ·����Ϊ���Կ��������ķ�չ��������ԣ����ǵķ���֧�������ĸ����������ȳ�����֢������-����Ⱥ����õ��Ŷ��ɴٽ��᳦���ķ�������֢�о��� 77��1���� 27-40�� ©2016 AACR��Dietary Emulsifier�CInduced Low-Grade Inflammation Promotes Colon Carcinogenesis | Cancer Research
https://cancerres.aacrjournals.org/content/77/1/27��
��֬��ʳͨ���ƻ���ϸ��Ĥ�����������ٳ���������
��֢Ԥ����־2016; 21��95-103
���߷�����2016��6��30��
©2016������֢Ԥ��ѧ�ᡣ
������
�����������Ӧ���ڰ�������Ƥ���ڵĸ�����֯���������ԵͶ���֢������������Ӧ���������о���Ŀ���ǵ����֬��ʳ��HFD����ApcMin / +С��ϸ��Ĥ�����Ժͳ�������������Ӱ�졣
������
��������ʳ��ND����HFDι��С��12�ܡ����㳦����������������ȷ���ڶ���Ѫ֢������Ӧ������֢�������־�ͨ����������ӫ���أ�FITC��-�Ͼ�������Ĥ��϶���ӵ��ױ����������������Եı仯��
�����
��ND����ȣ�HFD���������Ŀ���Ÿ��ߣ�P <0.05������ND����ȣ�HFD���ѪҺ�ܿ����������ϵͣ����������˵ı�־��᳦8-�ǻ�-2'-��������ˮƽ�ϸߣ�P <0.05���� HFD��FITC-���������������������ӣ�P <0.05������HFD��Ĥ��϶���ӵ��װ���zonula occludens-1��claudin-1��occludin�ı���ϵ͡�High-fat Diet Accelerates Intestinal Tumorigenesis Through Disrupting Intestinal Cell Membrane Integrity
Journal of Cancer Prevention 2016;21:95-103
Published online June 30, 2016
© 2016 Korean Society of Cancer Prevention.
Background:
Excess energy supply induces chronic low-grade inflammation in association with oxidative stress in various tissues including intestinal epithelium. The objective of this study was to investigate the effect of high-fat diet (HFD) on intestinal cell membrane integrity and intestinal tumorigenesis in ApcMin/+ mice.
Methods:
Mice were fed with either normal diet (ND) or HFD for 12 weeks. The number of intestinal tumors were counted and biomarkers of endotoxemia, oxidative stress, and inflammation were determined. Changes in intestinal integrity was measured by fluorescein isothiocyanate (FITC)-dextran penetration and membrane gap junction protein expression.
Results:
HFD group had significantly higher number of tumors compared to ND group (P < 0.05). Blood total antioxidant capacity was lower in HFD group, while colonic 8-hydroxy-2��-deoxyguanosine level, a marker of oxidative damage, was higher in HFD group compared to that of ND group (P < 0.05). The penetration of FITC-dextran was substantially increased in HFD group (P < 0.05) while the expressions of membrane gap junction proteins including zonula occludens-1, claudin-1, and occludin were lower in HFDHigh-fat Diet Accelerates Intestinal Tumorigenesis Through Disrupting Intestinal Cell Membrane Integrity
http://www.jcpjournal.org/journal/view.html?doi=10.15430/JCP.2016.21.2.95��
Article
Published: 07 May 2019
Lipopolysaccharide induces the differentiation of hepatic progenitor cells into myofibroblasts constitutes the hepatocarcinogenesis-associated microenvironment
Wen-ting Liu, Ying-ying Jing, Lu Gao, Rong Li, Xue Yang, Xiao-rong Pan, Yang Yang, Yan Meng, Xiao-juan Hou, Qiu-dong Zhao, Zhi-peng Han & Li-xin Wei
Cell Death & Differentiation volume 27, pages85�C101(2020)Cite this article
Abstract
Hepatocellular carcinoma (HCC) generally occurs in the presence of chronic liver injury, often as a sequela of liver fibrosis. Hepatic progenitor cells (HPCs) are known to be capable of forming both hepatocytes and cholangiocytes in chronic liver injury, which are also considered a source of myofibroblasts and tumor-initiating cells, under carcinogenic circumstances. However, the underlying mechanisms that activate HPCs to give rise to HCC are still unclear. In current study, the correlation between HPCs activation and liver fibrosis and carcinogenesis was investigated in rats and human specimens. We analyzed the role of HPCs in tumorigenesis, by transplanting exogenous HPCs in a diethylnitrosamine-induced rat HCC model. Our data indicated that HPC activation correlated with hepatic fibrosis and hepatocarcinogenesis. We further found that exogenous HPC infusion promoted liver fibrosis and hepatocarcinogenesis, while lipopolysaccharides (LPS) played an important role in this process. However, results of our study indicated that LPS did not induce HPCs to form tumor in nude mice directly. Rather, LPS induced myofibroblast-like morphology in HPCs, which enhanced the tumorigenic potential of HPCs. Further experiments showed that LPS/Toll-like receptor 4 (TLR4) signaling mediated the differentiation of HPCs into myofibroblasts and enhanced the production of interleukin-6 (IL-6) and tumor necrosis factor-�� (TNF-��), which led to the aberrant expression of Ras and p53 signaling pathways in HPCs, and finally, promoted the proliferation and malignant transformation of HPCs, by long non-coding RNA regulation. Besides, examination of HCC clinical samples demonstrated that IL-6 and TNF-�� production correlated with HPC activation, hepatic fibrosis, and HCC recurrence. Our study indicates that both myofibroblasts and tumor cells are derived from HPCs. HPC-derived myofibroblasts create tumor microenvironment and contribute to the proliferation and malignant transformation of HPCs. Furthermore, LPS present in the chronic liver inflammation microenvironment might play an important role in hepatocarcinogenesis, by regulating the plastic potential of HPCs.Lipopolysaccharide induces the differentiation of hepatic progenitor cells into myofibroblasts constitutes the hepatocarcinogenesis-associated microenvironment | Cell Death & Differentiation
https://www.nature.com/articles/s41418-019-0340-7��
Ferrichrome is a cyclic hexa- peptide that forms a complex with iron atoms. It is a siderophore composed of three glycine and three modified ornithine residues with hydroxamate groups [-N(OH)C(=O)C-]. The 6 oxygen atoms from the three hydroxamate groups bind Fe(III) in near perfect octahedral coordination. Ferrichrome was first isolated in 1952, has been found to be produced by fungi of the genera Aspergillus , Ustilago , and Penicillium . Ferrichrome - Wikipedia en.wikipedia.org/wiki/Ferrichrome
��
Probiotic-derived ferrichrome inhibits colon cancer progression via JNK-mediated apoptosis
1Division of Gastroenterology and Hematology/Oncology, Department of Medicine, Asahikawa Medical University, Asahikawa 078-8510, Japan
Previous reports have suggested that some probiotics inhibit tumorigenesis and cancer progression. However, the molecules involved have not yet been identified. Here, we show that the culture supernatant of Lactobacillus casei ATCC334 has a strong tumour-suppressive effect on colon cancer cells. Using mass spectrometry, we identify ferrichrome as a tumour-suppressive molecule produced by Lactobacillus casei ATCC334. The tumour-suppressive effect of ferrichrome is greater than that of cisplatin and 5-fluorouracil, and ferrichrome has less of an effect on non-cancerous intestinal cells than either of those agents. A transcriptome analysis reveals that ferrichrome treatment induces apoptosis, which is mediated by the activation of c-jun N-terminal kinase (JNK). Western blotting indicates that the induction of apoptosis by ferrichrome is reduced by the inhibition of the JNK signalling pathway. This we demonstrate that probiotic-derived ferrichrome exerts a tumour-suppressive effect via the JNK signalling pathway.
Probiotic-derived ferrichrome inhibits colon cancer progression via JNK-mediated apoptosis
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4987524/��
Lipopolysaccharide induces inflammation and facilitates lung metastasis in a breast cancer model via the prostaglandin E2-EP2 pathway
Inflammation is a potent promoter of tumor metastasis. The aim of the present study was to explore the function of systemic inflammation in the formation of lung metastasis of breast cancer cells in a mouse model. BALB/c mice were injected intraperitoneally with lipopolysaccharide (LPS) in order to establish an inflammatory animal model and 4T1 murine breast cancer cells were injected through the tail vein to induce lung metastasis. The levels of proinflammatory cytokines were evaluated by ELISA. Metastases on the surface of the lungs were counted and histologically analyzed by hematoxylin and eosin staining. Angiogenesis in the lungs was examined by CD31 immunofluorescence. Mouse pulmonary endothelial cells (MPVECs) were isolated and used to assay endothelial tube formation and determine the protein expression levels of vascular endothelial growth factor (VEGF) in vitro. Serum levels of VEGF and prostaglandin E2 (PGE2), the number and size of metastatic lesions, and the expression levels of cyclooxygenase‑2 were significantly greater in the lungs of LPS‑treated mice, as compared with those in control mice threated with phosphate‑buffered saline. Blood vessel density was also markedly increased in the LPS‑treated mice. These increases were reversed by treatment with celecoxib. In vitro, the protein expression levels of VEGF produced by the PGE2‑treated cells were significantly increased in a concentration‑dependent manner. In addition, the production of VEGF was increased in response to treatment with the PGE2 receptor (EP2) agonist ONO‑AE1‑259‑01; however, this increase was abrogated by treatment with AH6809, an EP2 receptor antagonist. Treatment with PGE2 or VEGF alone promoted the tube formation of MPVECs and this effect was reversed by treatment with celecoxib. These results demonstrated that PGE2 may regulate the release of VEGF by MPVECs through the EP2 receptor pathway and thereby promoted pulmonary angiogenesis and breast cancer metastasis in a mouse model.
Introduction��
Lipopolysaccharide induces inflammation and facilitates lung metastasis in a breast cancer model via the prostaglandin E2-EP2 pathway https://www.spandidos-publications.com/10.3892/mmr.2015.3258
��
Infection. 2018 Dec;46(6):751-760. doi: 10.1007/s15010-018-1178-5. Epub 2018 Jul 12.
Gut-origin sepsis in the critically ill patient: pathophysiology and treatment.
Assimakopoulos SF1, Triantos C2, Thomopoulos K2, Fligou F3, Maroulis I4, Marangos M5, Gogos CA5.
Author information 1Department of Internal Medicine, Division of Infectious Diseases, University of Patras Medical School, 26504, Patras, Greece. sassim@upatras.gr.2Department of Internal Medicine, Division of Gastroenterology, University of Patras Medical School, 26504, Patras, Greece.3Department of Anesthesiology and Critical Care Medicine, University of Patras Medical School, 26504, Patras, Greece.4Department of Surgery, University of Patras Medical School, 26504, Patras, Greece.5Department of Internal Medicine, Division of Infectious Diseases, University of Patras Medical School, 26504, Patras, Greece.
Abstract
INTRODUCTION: Gut permeability is increased in critically ill patients, and associated with the development of the systemic inflammatory response syndrome and multiple organ dysfunction syndrome (MODS). The pathogenetic link(s) and potential therapies are an area of intense research over the last decades.
METHODS: We thoroughly reviewed the literature on gut-origin sepsis and MODS in critically ill patients, with emphasis on the implicated pathophysiological mechanisms and therapeutic interventions.
FINDINGS:
Intestinal barrier failure leading to systemic bacterial translocation associated with MODS was the predominant pathophysiological theory for several years. However, clinical studies with critically ill patients failed to provide the evidence of systemic spread of gut-derived bacteria and/or their products as a cause of MODS. Newer experimental data highlight the role of the mesenteric lymph as a carrier of gut-derived danger-associated molecular patterns (DAMPs) to the lung and the systemic circulation. These substances are recognized by pattern recognition receptor-bearing cells in diverse tissues and promote proinflammatory pathways and the development MODS. Therefore, the gut becomes a pivotal proinflammatory organ, driving the systemic inflammatory response through DAMPs release in mesenteric lymph, without the need for systemic bacterial translocation.
CONCLUSIONS: There is an emerging need for application of sensitive non-invasive and easily measured biomarkers of early intestinal injury (e.g., citrulline, intestinal fatty acid protein, and zonulin) in our everyday clinical practice, guiding the early pharmacological intervention in critically ill patients to restore or prevent intestinal injury and improve their outcomes.
KEYWORDS: Bacterial translocation; Danger-associated molecular patterns; Gut-lymph hypothesis; Gut-origin sepsis; ICU; Intestinal barrier; Intestinal permeability
Gut-origin sepsis in the critically ill patient: pathophysiology and treatment. - PubMed - NCBI https://www.ncbi.nlm.nih.gov/pubmed/30003491
��
Crit Care Clin. 2016 Apr;32(2):203-12. doi: 10.1016/j.ccc.2015.11.004. Epub 2016 Feb 4.
The Gut as the Motor of Multiple Organ Dysfunction in Critical Illness.
Klingensmith NJ1, Coopersmith CM2.
Author information
1
Department of Surgery, Emory Critical Care Center, Emory University School of Medicine, Atlanta, GA, USA.
2
Department of Surgery, Emory Critical Care Center, Emory University School of Medicine, Atlanta, GA, USA. Electronic address: cmcoop3@emory.edu.
Abstract
All elements of the gut - the epithelium, the immune system, and the microbiome - are impacted by critical illness and can, in turn, propagate a pathologic host response leading to multiple organ dysfunction syndrome. Preclinical studies have demonstrated that this can occur by release of toxic gut-derived substances into the mesenteric lymph where they can cause distant damage. Further, intestinal integrity is compromised in critical illness with increases in apoptosis and permeability. There is also increasing recognition that microbes alter their behavior and can become virulent based upon host environmental cues. Gut failure is common in critically ill patients; however, therapeutics targeting the gut have proven to be challenging to implement at the bedside. Numerous strategies to manipulate the microbiome have recently been used with varying success in the ICU.The Gut as the Motor of Multiple Organ Dysfunction in Critical Illness. - PubMed - NCBI
https://www.ncbi.nlm.nih.gov/pubmed/27016162��
Autophagy
Taylor & Francis
Lipopolysaccharide mediates hepatic stellate cell activation by regulating autophagy and retinoic acid signaling
Ming Chen, Jiaxing Liu, [...], and Wenhua Ling
aDepartment of Nutrition, School of Public Health, Sun Yat-Sen University, Guangzhou, Guangdong, People's Republic of China
bGuangdong Provincial Key Laboratory of Food, Nutrition and Health, Guangzhou, Guangdong, China
ABSTRACT
Bacterial translocation and lipopolysaccharide (LPS) leakage occur at a very early stage of liver fibrosis in animal models. We studied the role of LPS in hepatic stellate cell (HSC) activation and the underlying mechanisms in vitro and in vivo. Herein, we demonstrated that LPS treatment led to a dramatic increase in autophagosome formation and autophagic flux in LX-2 cells and HSCs, which was mediated through the AKT-MTOR and AMPK-ULK1 pathway. LPS significantly decreased the lipid content, including the lipid droplet (LD) number and lipid staining area in HSCs; pretreatment with macroautophagy/autophagy inhibitors or silencing ATG5 attenuated this decrease. Furthermore, lipophagy was induced by LPS through the autophagy-lysosomal pathway in LX-2 cells and HSCs. Additionally, LPS-induced autophagy further reduced retinoic acid (RA) signaling, as demonstrated by a decrease in the intracellular RA level and Rar target genes, resulting in the downregulation of Bambi and promoting the sensitization of the HSC's fibrosis response to TGFB. Compared with CCl4 injection alone, CCl4 plus LPS injection exaggerated liver fibrosis in mice, as demonstrated by increased Col1a1 (collagen, type I, �� 1), Acta2, Tgfb and Timp1 mRNA expression, ACTA2/��-SMA and COL1A1 protein expression, and Sirius Red staining area, which could be attenuated by injection of an autophagy inhibitor. LPS also reduced lipid content in HSCs in vivo, with this change being attenuated by chloroquine (CQ) administration. In conclusion, LPS-induced autophagy resulted in LD loss, RA signaling dysfunction, and downregulation of the TGFB pseudoreceptor Bambi, thus sensitizing HSCs to TGFB signaling.
KEYWORDS: autophagy, BAMBI, hepatic stellate cell, liver fibrosis, LPS, retinoic acid signaling, TGFB��
INTRODUCTION
Liver fibrosis represents an adaptive response to repeated chronic liver injuries, which are primarily caused by chronic viral hepatitis and steatohepatitis associated with either alcohol consumption or obesity.1, 2 Studies using animal models of liver fibrosis have demonstrated that myofibroblasts, which are not present in normal liver tissue, play a crucial role in hepatic fibrogenesis. Myofibroblasts are derived from 3 cellular sources, liver resident hepatic stellate cells (HSCs), portal fibroblasts and bone marrow-derived collagen-producing cells, and the composition of myofibroblasts varies depending on the etiology of liver fibrosis.3 However, activated HSCs are the predominant type of myofibroblasts observed during the progression of liver fibrosis.4 Quiescent HSCs, which are located in the space of Disse and store retinoids in lipid droplets (LDs), transform into an activated phenotype characterized by the induction of fibrotic markers and depletion of LDs. However, the mechanisms underlying HSC activation remain unclear. Therefore, elucidating these HSC activation events could promote new strategies for preventing and treating liver fibrosis.
Autophagy is an evolutionarily conserved process in which cytoplasmic constituents, including damaged and dysfunctional proteins and organelles, are delivered to lysosomes for degradation and recycling of the breakdown products.5 Macroautophagy (hereafter referred to as autophagy) involves the formation of double-membraned vesicles called autophagosomes and the subsequent fusion of autophagosomes with lysosomes for cargo recycling to maintain cellular homeostasis.6 Defects in autophagy have been closely associated with many human diseases, including cancer, infection and neurodegeneration.7-9 Recent studies have demonstrated that functional autophagy is involved in lipid clearance in hepatocytes, a process termed lipophagy.10-12 Previous studies have shown that autophagy plays a critical role in lipid clearance and fibrotic activity,13-15 and this effect is associated with the liberation of free fatty acids from LDs and subsequent mitochondrial ��-oxidation, which provides energy to support HSC activation.16Bacterial lipopolysaccharide (LPS), a cell wall component of gram-negative bacteria that is among the strongest known inducers of inflammation, has been found to be associated with hepatic fibrogenesis through direct interactions with HSCs.17 During chronic liver injury caused by CCl4 injection or bile duct ligation (BDL), the plasma concentration of LPS is significantly elevated even at a very early stage due to changes in the intestinal mucosal permeability and increased bacterial translocation.17, 18 Consistent with these reports, one recent study observed that the transplantation of Gram-negative bacteria affects hepatic fibrogenesis after BDL.19 However, the molecular mechanism underlying the effects of LPS on HSC activation is poorly understood. Recent studies have shown that LPS induces autophagy in macrophages, linking 2 ancient processes, autophagy and innate immunity.20-22 Here, we hypothesized that the LPS regulates HSC activation through autophagy and retinoic acid signaling.
Results
LPS treatment induced autophagic markers in LX-2 cells
LPS exposure promoted autophagic flux in LX-2 cells and HSCs
The AKT-MTOR signaling pathway mediated LPS-induced autophagy
The AKT-MTOR cascade is considered a major negative regulator of autophagy in multiple types of cell.10, 23,24 Therefore, we examined whether this signaling pathway was also involved in LPS-induced autophagy. As shown in Fig. 2, LPS treatment led to significantly decreased phosphorylation of AKT at Thr308 and MTOR at Ser2448, and increased phosphorylation of AMPK at Thr172and ATG5 expression.
LPS stimulated HSC lipophagy
LPS mediated the downregulation of Bambi through autophagy and the subsequent promotion of the HSC fibrotic response to TGFB signaling
LPS-induced RA signaling dysfunction inhibited Bambi expression and promoted HSC activation
Autophagy is involved in the LPS-mediated augmentation of the fibrotic response in CCl4-treated mice
LPS induced autophagy and decreased RA signaling in HSCs in vivo
In summary, the present study demonstrates that LPS exposure promotes HSC fibrosis through increased autophagy activity and dysfunctional RA signaling. This novel mechanism underlying the LPS-induced fibrotic response of HSCs is associated with LDs loss and downregulation of the TGFB pseudoreceptor BAMBI.��
Lipopolysaccharide mediates hepatic stellate cell activation by regulating autophagy and retinoic acid signaling
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5788469/#__ffn_sectitle��
Probiotics for the airways: Potential to improve epithelial and immune homeostasis
K. Martens B. Pugin I. De Boeck I. Spacova B. Steelant S. F. Seys S. Lebeer P. W. Hellings
First published: 05 June 2018 https://doi.org/10.1111/all.13495 Citations: 12
The authors are supported by grants from the Belgian Federal Government (IUAP P7/30), the Institute for Science and Technology Flanders (IWT) (TBM project 130260), the research council of the KU Leuven (GOA 2009/07 and 14/011), and IWT�\SBO grant (ProCure, 150052).
Abstract
Probiotics are live microorganisms that, when administered in adequate amounts, confer health benefit on the host. The therapeutic effects of probiotics have been mostly studied in the gastrointestinal tract, but recent evidence points toward the potential of these bacteria to prevent and/or treat chronic airway diseases. In this review, possible mechanisms of action of probiotics in the airways are described, with a particular focus on their capacity to modulate the epithelial barrier function and their mode of interaction with the immune system. Indeed, probiotic bacteria, mostly lactobacilli, can promote the expression and regulation of tight junctions and adherence junctions, resulting in the restoration of a defective epithelial barrier. These bacteria interact with the epithelial barrier and immune cells through pattern recognition receptors, such as Toll�\like receptors, which upon activation can stimulate or suppress various immune responses. Finally, the clinical potential of probiotics to treat inflammatory diseases of the upper and lower respiratory tract, and the difference between their mode of application (eg, oral or nasal) are discussed here.
������������������Ƥ��������̬��DZ��
K.Martens B.Pugin I.De Boeck I.Spacova B.Steantant S.F.Seys S.Lebeer P.W.Hellings
�״η�����2018��6��5��https://doi.org/10.1111/all.13495���ô�����12
���ߵõ�����ʱ����������IUAP P7 / 30����������˹��ѧ�����о�Ժ��TBW��Ŀ130260����³���ѧ�о����»ᣨGOA 2009/07��14/011���������� ����IWT�\SBO���ProCure��150052����
����
�������ǻ����������㹻����ʹ��ʱ�����Ը�������������������������������Ҫ��θ�����н������о����������֤�ݱ�����Щϸ������Ԥ����/��������������������DZ��������ƪ�����У��������������������еĿ������û������ر��ע�������Ƥ���Ϲ��ܵ���������������ϵͳ����õķ�ʽ��ȷʵ��������ϸ������Ҫ����˾������Դٽ��������Ӻ�ճ�����ӵı���͵��ڣ��Ӷ��ָ���ȱ�ݵ���Ƥ���ϡ���Щϸ��ͨ��ģʽʶ�����壨����Toll�����壩����Ƥ���Ϻ�����ϸ������ã��������Դ̼������Ƹ������߷�Ӧ������������������������Ϻ��������º��������Լ������ٴ�DZ�����Լ����ǵ�Ӧ�÷�ʽ������ڷ��ǣ�֮��IJ��졣��
Abbreviations
AHR
Airway hyper�\reactivity
AJ
Adherence junction
AR
Allergic rhinitis
BALF
Bronchoalveolar lavage fluid
CRS
Chronic rhinosinusitis
DCs
Dendritic cells
IEC
Intestinal epithelial cell
IL
Interleukin
LAB
Lactic acid bacteria
LPS
Lipopolysaccharides
MAMPs
Microorganism�\associated molecular patterns
NGS
Next�\generation sequencing
NLRs
NOD�\like receptors
OVA
Ovalbumin
PRRs
Pattern recognition receptors
TGF�\��
Transforming growth factor ��
TJ
Tight junction
TLRs
Toll�\like receptors
TNF�\��
Tumor necrosis factor�\��
Tregs
Regulatory T cells
URT
Upper respiratory tract
ZO
Zonula occludens
1 INTRODUCTION
The human body is colonized by approximately 100 trillion microbes, collectively known as the microbiota, which play an essential role in our well�\being.1 Thanks to the development of advanced culture�\independent tools, such as next�\generation sequencing, insights have been obtained in possible associations of the microbiota composition with disease.2, 3 Together with additional experimental and clinical data on various roles of the microbiota, it is now widely accepted that the microbiota provides the host with a wide array of health effects. These health effects include (i) exclusion or inhibitions of pathogens,4, 5 (ii) enhancement of epithelial barrier function by the modulation of signaling pathways,6, 7 and (iii) strain�\specific modulation of local and systemic host immune responses (Figure 1).8
Figure 1
Open in figure viewerPowerPoint
Relationship between microbiota diversity, epithelial Integrity, and inflammation in disease state
The clinical potential of probiotics has gained interest as a novel oral and/or nasal treatment option for chronic airway diseases. Probiotics interact, via their microorganism�\associated molecular patterns (MAMPs), with pattern recognition receptors (PRRs) on epithelial cells. This interaction can modulate tight junctions (TJs) and adherence junctions (AJs), which results in the restoration of a defective epithelial barrier. Probiotics are able to modulate host immune responses via interactions with dendritic cells (DCs.), which on their turn promote regulatory T cells (Tregs) and reduce T helper 1 (Th1) and T helper 2 (Th2) cells.
The involvement of the microbiota in the development or exacerbation of chronic noninfectious diseases is also increasingly considered.9 An imbalance of the microbiota composition, also known as dysbiosis, has been shown in various studies to negatively impact the health status.10 Three subtypes of dysbiosis have been identified as follows: (i) loss of beneficial microbial agents, (ii) expansion of potentially harmful microorganisms, and (iii) loss of overall microbial diversity.10 Microbial dysbiosis has been identified in different chronic inflammatory diseases, including chronic rhinosinusitis (CRS),11, 12 asthma,13, 14 Crohn's disease,15 and ulcerative colitis.16 Interestingly, in these chronic diseases, a disturbed permeability and function of the epithelial barrier have also been demonstrated.17-19 It is, however, not known whether microbiota dysbiosis could be the cause or simply a consequence of the associated epithelial barrier disruption.
When looking specifically at the airways, various microorganisms such as, Staphylococcus aureus,20 Streptococcus pneumoniae, and Haemophilus influenzae21 could induce pathogenic effects on the host, at least in part by increasing the epithelial barrier permeability in both the upper and lower airways. On the contrary, other microorganisms��both endogenous and exogenous��can also provide beneficial effects22 (Figure 2). These beneficial bacteria are called probiotics, defined as live microorganisms that, upon administration in adequate amounts, can provide health benefits to the host.23 Traditionally, Lactobacillus and Bifidobacterium strains have been used as probiotics, mainly to reduce gastrointestinal tract discomfort. Other bacterial taxa have probiotic potentials, such as the Streptococcus taxa24 or Dolosigranulum which is associated with respiratory health in different microbiome studies,25, 26 but these studies are limited so that this review will mainly focus on the Lactobacillus strains.
image
Figure 2
Open in figure viewerPowerPoint
Interaction of probiotics with the epithelial barrier and the immune system. Probiotics induce their health�\promoting capacities via different mechanisms. These bacteria are able to enhance epithelial barrier function via modulation of the intercellular junctions (TJs, AJs, and desmosomes) or via interaction with the different PRRs, present on the epithelial barrier. These PRRs (ie, TLRs, CLRs, NLRs) recognize the MAMPs, such as LTA, CPS, and LPS, present on the probiotic cell surface. Probiotics are also able to modulate the local and systemic immune responses. They have the ability to interact with DCs, present between the epithelial cells or in the submucosal region. This interaction can result in the activation of Tregs. Additionally, it has been suggested that Tregs play a role in maintaining epithelial barrier via the production of TGF�\�� and IL�\10. Finally, probiotics are able to modulate the Th1 and Th2 responses, resulting in the restoration of the immune homeostasis. DCs, dendritic cells; Tregs, regulatory T cells; PRRs, pattern recognition receptors; TLRs, Toll�\like receptors; CLRs, C�\type lectin receptors; NLRs, NOD�\like receptors; LTA, lipoteichoic acid; CPS, cell wall�Cassociated polysaccharide; LPS, lipopolysaccharide
Interestingly, lactobacilli were shown to be normal inhabitants of the healthy nasopharynx and upper respiratory tract (URT).27-29 For instance, Bogaert et al27 showed a predominance of Lactobacillus species in children in summer (absolute abundance is 96%) compared to winter (absolute abundance is 10%). Additionally, lactobacilli were also detected in the healthy nasopharynx of Chinese individuals30 and in the nasopharynx of healthy Belgian individuals (up to 10% relative abundance).31 Lactobacilli are also found in the tonsillar crypts of both healthy adults and infants.32 Nevertheless, whether lactobacilli could also function as probiotics in the nasopharyngeal niche is currently underexplored.
In this review, we summarize documented beneficial effects of probiotic microorganisms (mainly Lactobacillus and other species) and discuss their potential mechanisms of action in chronic diseases of the upper and lower airways. As the studies of the gastrointestinal impact of probiotics are more advanced, some data from this field are incorporated as additional supporting evidence. We describe documented barrier�\restoring capacities of probiotics, their mechanisms of interaction with the innate immune system, and the potential clinical applications of these bacteria to treat different chronic inflammatory diseases of the upper URT.
2 PROBIOTIC REGULATION OF THE EPITHELIAL BARRIER FUNCTION
The epithelial barrier is the first physical barrier protecting the human body from the entrance of harmful substances including allergens, pathogens, and irritants.33 Intercellular junctions, such as tight junctions (TJs), adherence junctions (AJs), and desmosomes, play a critical role in the formation and maintenance of the physical barrier. TJs are the most apically located junctions and provide the epithelia with a semipermeable size�\ and ion�\specific barrier, restricting diffusion of macromolecular components.18, 34, 35 AJs are positioned immediately below TJs and are essential for cell�\cell adhesion, regulation of the actin cytoskeleton, intracellular signaling, and transcriptional regulation in epithelial cells.36 Desmosomes form the third intercellular junction complex and are important in maintaining barrier integrity and cell adhesion.37
The beneficial effects of probiotics on epithelial barrier dysfunction have been extensively studied in the gastrointestinal tract (Table 1). For instance, Lactobacillus plantarum MB452 has been shown to increase the expression levels of TJ�\related genes in healthy intestinal epithelial cells (IECs) in vitro.38 Similar positive effects on intestinal epithelial barrier integrity and TJ expression were observed with other probiotic strains such as Lactobacillus rhamnosus GG,39 Streptococcus thermophiles ATCC19258, L. plantarum MB452,40 and the Gram�\negative probiotic strain Escherichia coli Nissle 1917.41, 42 Additionally, certain Lactobacillus strains appear to have the capability to increase epithelial barrier integrity through the stabilization of AJs expression.43 More specifically, the lactobacilli tested could increase the expression of E�\cadherin and �©\catenin and decrease the abundance of protein kinase C (enzyme involved in the disassembling of AJs) in T84 human colonic epithelial cell line.43 Various barrier�\restoring properties of probiotics have also been demonstrated in different in vivo models. For instance, in a murine model of colitis induced by dinitrobenzene sulfonic acid, oral administration of L. rhamnosus CNCM I�\3690 protected epithelial barrier integrity by modulating TJ (occludin) and AJ molecules (E�\cadherin).44
Table 1. Barrier�\restoring capacities of different probiotics in the gastrointestinal tract
Strain TJ or AJ proteins Cells or mouse model Barrier function Ref.
L. plantarum MB452 �� OCCL, ZO�\1, ZO�\2, and cingulin expression Caco�\2 cells Increase in barrier integrity 38
L. rhamnosus GG �� ZO�\1, claudin�\1, and OCCL expression Caco�\2 cells Restoration of gliadin induced epithelial barrier disruption 39
S. thermophiles ATCC19258 and L. acidophilus ATCC4356 Maintenance of actin and ZO�\1, �� actinin and OCCL expression HT29/cl.19A and Caco�\2 cells Increase in barrier function of untreated epithelial cells 40
E. coli Nissle 1917 �� ZO�\1 expression Human small and large intestinal epithelial cells and DSS�\treated mice Upregulation of TJ expression in murine IECs and protection against DSS colitis induced increase in mucosal permeability 41
E. coli Nissle 1917 �� ZO�\2 expression, silencing of PKC�� T84 cells Restoration of a disrupted epithelial barrier 42
L. acidophilus, L. fermentum, L. gasseri and L. rhamnosus �� Stabilization of E�\cadherin and �©\catenin T84 cell monolayers Increase in barrier integrity 43
L. rhamnosus CNCM I�\3690 �� OCCLN and E�\cadherin expression Caco�\2 cells and murine model of colitis induced by DNBS Restoration of intestinal barrier function and increased levels of TJ and AJ proteins 44
In the airways, there are currently limited reports describing the regulatory properties of probiotics on the epithelial barrier. In vivo, it has been shown that oral treatment with L. rhamnosus CRL1505 could prevent the polycytidylic acid [poly (I:C)]�\induced increased permeability of the bronchoalveolar�\capillarity barrier, as determined by albumin levels in the lungs.45 This improvement was linked to a decrease in the activation of pro�\inflammatory cells and the production of pro�\inflammatory cytokines in the lungs.45 Similar findings were observed using nasally administered Lactococcus lactis NZ9000, which could counteract S. pneumoniae�\induced lung tissue permeability.46 In vitro, the stimulation of Calu�\3 bronchial epithelial cells with the synthetic bacterial lipopeptide Pam3CysSK4 resulted in a concentration�\dependent increase in epithelial barrier function and decrease in epithelial permeability, as a result of an increased expression of the TJ proteins claudin�\1 and Zonula occludens (ZO) �\1 and a decreased expression of occludin.47
3 PROBIOTIC INTERACTIONS WITH THE IMMUNE SYSTEM
3.1 Interaction with pattern recognition receptors
Probiotics can modulate local and systemic host immune responses in a strain�\specific manner via the expression of different microorganism�\associated molecular patterns (MAMPs), such as lipopolysaccharides (LPS), flagellin, CpG�\DNA, and other surface (lipo)proteins.5 MAMPs are recognized by pattern recognition receptors (PRRs) present on epithelial and innate immune cells (Figure 2, Table 2). The best�\studied PRRs are the Toll�\like receptors (TLRs), NOD�\like receptors (NLRs), and C�\lectin type receptors.48 Thanks to the broad specificity of PRRs, a limited number of them can detect a wide range of MAMPs.48 Interaction between MAMPs and PRRs leads to the activation of multiple signaling cascades that generates an appropriate molecular response against the encountered microorganism.49, 50 In a 56�\day murine ovalbumin (OVA)�\induced asthma model, it has been demonstrated that oral treatment with L. rhamnosus NutRes1 and Bifidobacterium breve M�\46 was able to modulate the mRNA expression levels of Tlr9 and Tlr3.51 However, these results need to be analyzed with prudence, because the increase in mRNA expression levels of Tlr3 and Tlr9 does not necessarily prove that these 2 probiotics were recognized directly by the receptors. A similar reduction in the type 1 immune response and injury in the lungs were observed in a murine model of sepsis where treatment with L. rhamnosus GG and Bifidobacterium longum (strain not specified) via oral gavaging could attenuate neutrophil infiltration in the lungs and decreased the expression of interleukin (IL)�\6 and tumor necrosis factor�\�� (TNF�\��). The beneficial effects of these probiotics appeared to be associated with decreased gene expression levels of TLR2 and MyD88 in the lungs of the mice.52 Finally, oral treatment with Lactobacillus reuteri ATCC 23272 of OVA�\sensitized mice has been shown to result in attenuation of inflammatory cell influx to the lungs and airway hyper�\reactivity (AHR). These beneficial effects seemed to be dependent on TLR9, because the treatment of TLR9−/− mice with this L. reuteri strain did not attenuate inflammatory cell influx in the lungs or AHR. Additionally, the attenuation of inflammatory cell influx was associated with reduced levels of pro�\inflammatory cytokines such as IL�\5 and IL�\13 in the bronchoalveolar lavage fluid (BALF) of the mice. However, L. reuteri ATCC 23272 could not alter Th1�\type cytokines IL�\12 and IFN�\�� or the anti�\inflammatory cytokine IL�\10 in BALF.53 Moreover, the authors showed that these effects were strain�\specific, because oral treatment with Lactobacillus salivarius AH102 did not improve allergic airway response in the OVA�\sensitized mice.53 This is in agreement with the fact that the sum of all MAMPs expressed by a certain probiotic is strain�\specific, resulting in their differential effects on epithelial and/or immune cells.5, 54
Table 2. Examples of interactions of probiotics with the immune system (relevant for respiratory allergies)
Strain Mechanism Outcome Ref.
L. rhamnosus NutRes1 and Bifidobacterium breve M�\46 Reduction of chronic allergic inflammation by increasing the mRNA expression levels of Tlr9 and Tlr3 Suppression of pulmonary airway inflammation, induction of airway remodeling and inhibition of mast cell degranulation (48)
L. rhamnosus GG and Bifidobacterium longum ATCC Attenuation of neutrophil infiltration in the lungs and decreased expression of IL�\6 and TNF�\�� via decreased gene expression of TLR2 and MyD88 Reduction of immune response and lung injury (49)
L. reuteri ATCC 23272 Modulation of TLR9 resulted in reduced levels of pro�\inflammatory cytokines such as IL�\5 and IL�\13 Attenuation of inflammatory cell influx to the lungs and AHR (50)
L. reuteri and L. casei Modulation of DC function through dendritic cell�\specific intercellular adhesion molecule 3�\grabbing non�\integrin in vitro Induction of IL�\10 producing Tregs in vitro (67)
L. rhamnosus GG and B. lactis Bb�\12 Induction of Tregs, associated with increased expression of TGF�\�� in vivo Inhibition of allergic sensitization and airway disease in an asthma mouse model (64)
L. casei IBSO41 and L. acidophilus D031 Stimulation of DCs in vitro Increased production of TGF�\�� (69)
L. plantarum NCIMB8826 Inhibition of specific IgE response, high levels of specific IgG2a and increased production of IFN�\�� Inhibition of an allergic Th2 profile (63)
L. rhamnosus GG Significant decrease in eosinophils, IL�\13 and IL�\5 levels in the lungs, and airway hyperactivity Prevention of birch�\pollen�\induced allergic asthma by LGG (62)
To our knowledge, there is currently no in vitro evidence showing the interaction between probiotics and PRRs using primary nasal or bronchoepithelial cells. Nonetheless, the modulating effects of TLRs by probiotics have been investigated using intestinal cell lines. For example, Ganguli et al55 have shown the anti�\inflammatory effects of L. rhamnosus GG and its SpaC pilus adhesin through the modulation of the TLR/IL�\1R signaling pathway. The investigators demonstrated that T84 cells stimulated with Salmonella enterica serovar Typhimurium SL1344 or LPS can reduce IL�\1�� and IL�\6 secretion when treated with L. rhamnosus GG. These effects were due to the binding of SpaC pilus adhesin to the IECs, resulting in the downregulation of the mRNA levels of TLR3 and TLR4.55 In addition, phenotypic comparison of wild�\type and pili mutant strain of L. rhamnosus GG showed that the presence of pili reduced IL�\8 induction in Caco�\2 cells, which was at least partially mediated by TLR2.56 As the innate immune cells expressing PRRs provide a link between the innate and adaptive immunity, the described effects can further translate into the modulation of different T�\cell subsets and the corresponding cytokines.
3.2 Modulation of Th1/Th2 and regulatory T�\cell immune response by probiotics
Probiotic applications have been shown to impact effector and regulatory T�\cell responses in airway disease, resulting in the restoration of the Th1/Th2 immune balance,57 and/or stimulation of regulatory cytokine production and the regulatory T�\cell (Treg) subset.58-61 This typically leads to a reduction in lung Th2 inflammatory cytokine levels (eg, IL�\5 and IL�\13) and the attenuation of AHR, as recently demonstrated for preventive intranasal administration of L. rhamnosus GG in a mouse model of birch pollen�Cinduced allergic asthma.62 Additional immunomodulatory properties of probiotics in airway disease, including the stimulation of mucosal IgA levels and allergen�\specific B�\ and T�\cell responses, are well described elsewhere.63
Allergic airway disease is typically a result of excessive Th2 reactions; therefore, the ability of probiotic strains to stimulate a counterbalancing Th1 immune response can be a valuable mechanism of airway disease prevention and treatment. In vitro, co�\incubation with L. plantarum NCIMB885 induced a Th1�\biased cytokine profile in murine dendritic cells (DCs), which could be linked to lipoteichoic acid composition of this probiotic strain.64 In vivo, orally administered L. rhamnosus GG led to an increase in Th1 (IL�\12 and IFN�\��) and a decrease in Th2 (IL�\4, IL�\5, and IL�\13) BALF cytokine levels in a mouse model of OVA�\induced airway inflammation.65 This led to a reduction in inflammatory cell counts in the BALF (ie, eosinophils, lymphocytes, and monocytes) and AHR.65 The development of a Th1 over Th2 response was likewise observed as a result of intranasal co�\application of L. plantarum NCIMB8826 and the clinically relevant house dust mite aeroallergen Der p 1 in a mouse model of Der p 1 sensitization.66 This was reflected in an increase in specific IgG2a levels, a decrease in IFN�\�� production by restimulated splenocytes, and an inhibition of specific IgE production. These effects were accompanied by the attenuation of airway inflammation after house dust mite extract challenge, as demonstrated by reduced BALF eosinophil counts.66
Marked improvement of airway disease symptoms in murine models as a result of probiotic administration has also been linked to the induction of Tregs.67, 68 Tregs are a subset of T lymphocytes, pivotal in maintaining immune homeostasis through the suppression of self�\reactive T�\cell activation and expression.69 Their function is tightly linked to the regulatory IL�\10 and transforming growth factor (TGF)�\�� cytokines, which can drive the activation and proliferation of Tregs along with their suppressive functions. These regulatory cytokines can be produced by DCs58-60 or T cells61 in response to interactions with probiotic strains. For example, L. reuteri ASM20016 and Lactobacillus casei NIZO B255 have been shown to bind DC�\SIGN on DCs, which in turn drove the development of IL�\10�\producing Tregs.70 Also, for L. rhamnosus GG, it was recently shown that glycosylated heteropolymeric pili are involved in such DC�\SIGN�\mediated interactions with DCs.71 In another study, Bifidobacterium lactis AD011 increased the production of IL�\10 in a co�\culture with DCs and IECs, whereas L. casei IBS041 and Lactobacillus acidophilus AD031 increased TGF�\�� secretion in this co�\culture system.72 Similar results were obtained in another co�\culture system containing peripheral blood mononuclear cells and L. lactis, that is, an increased secretion of the anti�\inflammatory cytokine IL�\10.73 The in vitro data are further supported by in vivo studies in mouse models of allergic airway disease. Feleszko et al67 have shown that orally administered L. rhamnosus GG or B. lactis Bb�\12 can inhibit subsequent allergic sensitization and airway disease symptoms in a murine model of OVA�\induced asthma. These effects are partially due to the induction of Foxp3�\expressing Treg cells in the peribronchial lymph nodes, as well as increased TGF�\�� production by T cells in the mesenteric lymph nodes67. Likewise, oral treatment with L. rhamnosus NutRes1 and B. breve M�\46 stimulated Foxp3 expression in blood Treg cells and decreased the levels of various pro�\inflammatory cytokines such as IL�\6 and IL�\4 in the thoracic lymph nodes in a murine OVA�\induced asthma model.51 This resulted in beneficial effects to the same extent as the corticosteroid budesonide, that is, suppression of pulmonary airway inflammation, the induction of airway remodeling, and the inhibition of mast cell degranulation.74 Also, intranasal application of probiotic strains, such as L. paracasei NCC2461, has been shown to increase lung CD4+CD25+FoxP3+ Treg cell counts in a mouse model of allergic airway inflammation and to attenuate the allergic response.75
Taken together, these studies emphasize the ability of certain probiotic strains to shift the immune balance away from pathological responses, suggesting their potential application for prevention and/or treatment of airway inflammatory diseases.
4 CLINICAL APPLICATION OF PROBIOTICS FOR THE AIRWAYS
The beneficial effects of probiotics are highly influenced by different parameters such as duration and route of administration as well as the type of strain. Various studies have used murine models of allergic airway diseases to investigate the effects of probiotics. The administration route of these probiotics is typically oral, via drinking water or food,76-78 or through intragastric intubation.75 More recently, intranasal administration of probiotic bacteria has been increasingly explored in different mice studies.62, 75, 79 In the study of Spacova et al, the authors demonstrated that intranasal administration of L. rhamnosus GG attenuated birch pollen�Cinduced asthma in mice.62 However, when these mice were intragastric pretreated with L. rhamnosus GG, there was no significant effect on allergic sensitization.62 The effectiveness of intranasal application was also demonstrated in the study of Pellaton et al75 Their findings demonstrated that intranasal application of L. paracasei NCC2461 can modify both the systemic and local immune response and allows for niche�\specific effects compared to oral treatment. Also, clinical trials in humans to investigate the effects of probiotics on the airways are recently booming.80 In most clinical trials, the probiotics are orally administered, for instance via capsules or supplemented in dairy products, and exert their effects mostly via modulation of the immune system. However, based on the results obtained in different murine models, we believe that local administration (eg, nasal spray) can induce more niche�\specific effects both in the upper and in the lower airways, leading to a more effective treatment.
4.1 Probiotics for the upper respiratory tract diseases
Different randomized placebo�\controlled trials have been performed to investigate the potential of oral administration of probiotics in upper airway diseases.80, 81 In the meta�\analysis of Peng et al82, authors analyzed 11 randomized controlled trials in which probiotic efficiency for the prevention and/or treatment of allergic rhinitis (AR) was investigated. Results showed that in these 11 trials, probiotic treatment resulted in the improvement of the quality of life and nasal symptoms of the patients. However, none of the probiotic strategies tested could prevent AR nor improve the inflammatory markers in these patients. In the more recent meta�\analysis of Guvenc et al,81 22 randomized double�\blind placebo�\controlled studies were included, of which 17 studies demonstrated an improvement of the clinical symptoms in AR patients such as nasal blockage, rhinorrhea, and nasal itching. These beneficial effects were in 8 of the included trials linked with a decrease in inflammatory markers.81 The combination of different strains has also been studied in patients with AR. For instance, Koyama and colleagues tested a probiotic yoghurt containing a mixture of L. rhamnosus GR1 and Bifidobacterium adolescentis in 36 subjects with AR. They found no significant effects on clinical outcomes of the participants, but did observe increased serum levels of IL�\10 and IL�\12 during grass pollen season, and increased TGF�\�� in ragweed season in the probiotic group compared to the placebo group.83 Thus, although no effects on the patients' symptoms were observed, important regulatory markers were increased in these patients after probiotic administration.
Although the above�\mentioned studies indicate that probiotics may improve the overall quality of life and nasal symptoms of AR patients, strong evidence is still lacking to substantiate preventive and/or therapeutic roles for probiotics in AR. Moreover, nasal application of probiotics has not yet��to the best of our knowledge��been tested in AR patients. Nasal application of probiotics can lead to the modulation of inflammation in the lower airways which was shown in a mouse model of allergic airway inflammation.75 These results are very promising because asthma is an important comorbidity of AR.84
In contrast to AR, the use of probiotics in patients with CRS is even less explored. In a small�\scale pilot study, patients with CRS were shown to have reduced bacterial diversity with specific depletion of beneficial lactobacilli in brush samples of mucosal surfaces of the lateral, central, and medial portions of the maxillary sinuses.85 Interestingly, these same authors could subsequently show that Lactobacillus sakei ATCC 15521 was sufficient to protect against Corynebacterium tuberculostearicum�\induced sinusitis after intranasal inoculation in mice.85 The authors focused on L. sakei because they found this species in higher abundances in the mucosa of healthy subjects and it was among the taxa that were significantly depleted in CRS patients.85 This was one of the first studies to demonstrate that specific enrichment of the URT with lactobacilli can result in significant health benefits. In humans, a randomized placebo�\controlled trial in CRS patients that were treated orally for 4 weeks with L. rhamnosus R0011 showed no improvement in sinonasal quality of life scores (SNOT�\20 questionnaire).86 Another double�\blind placebo�\controlled multicenter study demonstrated that oral application of Enterococcus faecalis Symbioflor 1 for 6 months could result in a reduction in the frequency of acute exacerbations of CRS. Furthermore, these beneficial effects were sustained for 8 months after the end of the treatment.87 In line with these results, Kitz et al have reported a reduction in frequency and duration of recurrent acute sinusitis in children treated orally with E. faecalis Symbioflor�\1 for 8 weeks.88
To the best of our knowledge, only 1 placebo�\controlled trial has investigated the potential of a nasal spray for the treatment of CRS without nasal polyps.79 In this trial, 20 patients received a nasal spray containing 13 honeybee lactic acid bacteria (LAB) species (nine Lactobacillus and 4 Bifidobacterium species) for 2 weeks. After 2 weeks, the symptoms of the patients (SNOT�\22 questionnaire) were evaluated, together with inflammatory markers in nasal lavage fluid and their microbiome. This study proved that the nasal LAB spray was well tolerated by the patients without any side effects.79 However, there was no significant improvement with these honeybee isolates in symptom severity, nor changes in the immune response or microbiome of the patients.79 Nevertheless, this study is a first important step toward the use of nasal probiotic sprays for the treatment of CRS. Also, other studies exploring the use of a nasal spray containing probiotics for other URT infections, such as otitis media, already reported the safety of a probiotic spray in the nasal cavity.24, 89, 90 Additionally, we believe that better strain selection (especially human isolates from the URT) might be needed to observe significant effects on the clinical symptoms of patients.
4.2 Probiotics for the lower respiratory tract diseases
When studying the treatment potential of probiotics in chronic inflammatory diseases of the URT (eg, AR and CRS), it is important to consider that these diseases are often associated with comorbidities in the lower airways, that is, mainly asthma.84 Clinical studies investigating the potential of probiotics to treat patients with asthma are preliminary but show some promising results. For instance, Chen et al performed a randomized, double�\blind, placebo�\controlled trail to investigate the clinical potential of Lactobacillus gasseri PM�\A0005 in children (6�\12 years old) with asthma.91 After 2 months of daily treatment with 2x109 CFU/capsule of L. gasseri PM�\A0005, they measured an improvement in asthma symptom score, together with an improvement in airway functions, that is, peak expiratory flow rates.91 Additionally, they observed a significant reduction in the blood levels of pro�\inflammatory cytokines in the patients, such as TNF�\a, IFN�\��, IL�\12, and IL�\13.91 In line with these results, Gutkowski et al showed that oral treatment with Trilac, a mixture containing Lactobacillus acidophilus, Lactobacillus delbruecki ssp. bulgaricus, and Bifidobacterium bifidum, for 12 weeks in children with mild�\to�\moderate atopic asthma, resulted in an improvement in lung function and a reduction in exacerbations of asthma.92 In another study, van de Pol et al investigated the effect of synbiotic treatment in patients with asthma and house dust mite allergy.93 They demonstrated that treatment with the combination of B. breve M�\16V and galacto�\ and fructo�\oligosaccharide for 4 weeks had no effect on allergen�\induced changes in sputum eosinophils or lung function in these patients. However, a significant improvement in the baseline lung function and IL�\5 levels in the serum was observed.93 Finally, in the study of Rose et al, the authors performed a placebo�\controlled clinical study where they investigated 6 months of L. rhamnosus GG food supplementation in infants with atopic predisposition and recurrent wheezing.94 The results demonstrated no improvement in clinical outcomes, measured via severity scoring of atopic dermatitis and asthma symptom score (Ref Rose et al 2010). However, oral treatment with L. rhamnosus GG resulted in mild reduction in sensitization to aeroallergens as predictor for bronchial asthma.94 More information regarding the involvement of airway microbial dysbiosis in asthma and the potential of probiotics to treat these patients are extensively reviewed elsewhere.95
Although these studies are promising, more research is needed to evaluate whether targeting the URT via nasal and/or oral probiotic treatment is efficient enough to modulate the lung microbiota.
5 CONCLUDING REMARKS
As the first culture�\independent report on the presence of a healthy lung microbiota,96 studies demonstrated the complex microbial communities residing in both the upper and the lower airways of healthy individuals.97, 98 A dysbiosis of the microbiota has been shown in chronic airway diseases which, interestingly, seems to occur concomitantly with a defective epithelial barrier, as observed in asthma,19 AR,18, 99 and CRS.17 Therefore, we believe that probiotics will represent a novel treatment option for chronic airway diseases. Probiotics can modulate nasal epithelial barrier via modulation of TJs and AJs and interact with the innate immune system, resulting in the restoration of mucosal barrier integrity.
Furthermore, the nature and extent of these beneficial effects might vary depending on the probiotic strain and administration route. Recent studies point toward the niche�\specific effect of nasal administration of probiotics. Further research is nevertheless warranted to investigate the potential of nasal administration for the treatment of upper and lower airway diseases.
In addition, most of the results are obtained using in vitro and in vivo models of the intestine, and therefore, further research is needed to prove the beneficial effects of probiotics in the upper and lower airways.��
Probiotics for the airways: Potential to improve epithelial and immune homeostasis - Martens - 2018 - Allergy - Wiley Online Library
https://onlinelibrary.wiley.com/doi/full/10.1111/all.13495��
The American Journal of Pathology
Volume 187, Issue 3, March 2017, Pages 476-486
The American Journal of Pathology
Changes in the Luminal Environment of the Colonic Epithelial Cells and Physiopathological Consequences
Author links open overlay panelFrançoisBlachier∗MartinBeaumont∗MireilleAndriamihaja∗Anne-MarieDavila∗AnnaïgLan∗MartaGrauso∗LucieArmand∗RobertBenamouzig∗†DanielTom��∗
UMR Physiologie de la Nutrition et du Comportement Alimentaire, AgroParisTech, INRA, Universit�� Paris-Saclay, Paris, France
†
Department of Gastroenterology, Avicenne Hospital, Assistance Publique-Hôpitaux de Paris, Bobigny, France
��
Evidence, mostly from experimental models, has accumulated, indicating that modifications of bacterial metabolite concentrations in the large intestine luminal content, notably after changes in the dietary composition, may have important beneficial or deleterious consequences for the colonic epithelial cell metabolism and physiology in terms of mitochondrial energy metabolism, reactive oxygen species production, gene expression, DNA integrity, proliferation, and viability.Recent data suggest that for some bacterial metabolites, like hydrogen sulfide and butyrate, the extent of their oxidation in colonocytes affects their capacity to modulate gene expression in these cells. Modifications of the luminal bacterial metabolite concentrations may, in addition, affect the colonic pH and osmolarity, which are known to affect colonocyte biology per se. Although the colonic epithelium appears able to face, up to some extent, changes in its luminal environment, notably by developing a metabolic adaptive response, some of these modifications may likely affect the homeostatic process of colonic epithelium renewal and the epithelial barrier function. The contribution of major changes in the colonocyte luminal environment in pathological processes, like mucosal inflammation, preneoplasia, and neoplasia, although suggested by several studies, remains to be precisely evaluated, particularly in a long-term perspective.
Changes in the Luminal Environment of the Colonic Epithelial Cells and Physiopathological Consequences - ScienceDirect
https://www.sciencedirect.com/science/article/pii/S0002944016305429��
Published: 14 July 2017
(Dis)Trust your gut: the gut microbiome in age-related inflammation, health, and disease
Thomas W. Buford
Microbiome volume 5, Article number: 80 (2017) Cite this articleDepartment of Medicine, University of Alabama at Birmingham
Abstract
Chronic inflammation represents one of the most consistent biologic features of aging. However, the precise etiology of persistent low-grade increases in inflammation remains unclear. Recent evidence suggests that the gut microbiome may play a key role in age-related inflammation. Indeed, several studies have indicated that older adults display an altered composition of the gut microbiota, and early evidence indicates that this dysbiosis is associated with the presence of several key circulating inflammatory analytes. The present review summarizes knowledge on age-related inflammation and discusses how potential relationships with gut dysbiosis may lead to novel treatment strategies in the future.
��The pattern of disease is an expression of the response of man to his total environment (physical, biological, and social); this response is, therefore, determined by anything that affects man himself or his environment.�� �C Rene Dubos, 1961(Dis)Trust your gut: the gut microbiome in age-related inflammation, health, and disease | Microbiome | Full Text
https://microbiomejournal.biomedcentral.com/articles/10.1186/s40168-017-0296-0��
Alzheimers Res Ther. 2018; 10: 124.
The gut microbiota-derived metabolite trimethylamine N-oxide is elevated in Alzheimer��s disease
Background
Trimethylamine N-oxide (TMAO), a small molecule produced by the metaorganismal metabolism of dietary choline, has been implicated in human disease pathogenesis, including known risk factors for Alzheimer��s disease (AD), such as metabolic, cardiovascular, and cerebrovascular disease.
Methods
In this study, we tested whether TMAO is linked to AD by examining TMAO levels in cerebrospinal fluid (CSF) collected from a large sample (n = 410) of individuals with Alzheimer��s clinical syndrome (n = 40), individuals with mild cognitive impairment (MCI) (n = 35), and cognitively-unimpaired individuals (n = 335). Linear regression analyses were used to determine differences in CSF TMAO between groups (controlling for age, sex, and APOE ��4 genotype), as well as to determine relationships between CSF TMAO and CSF biomarkers of AD (phosphorylated tau and beta-amyloid) and neuronal degeneration (total tau, neurogranin, and neurofilament light chain protein).
Results
CSF TMAO is higher in individuals with MCI and AD dementia compared to cognitively-unimpaired individuals, and elevated CSF TMAO is associated with biomarkers of AD pathology (phosphorylated tau and phosphorylated tau/A��42) and neuronal degeneration (total tau and neurofilament light chain protein).
Conclusions
These findings provide additional insight into gut microbial involvement in AD and add to the growing understanding of the gut�Cbrain axis.The gut microbiota-derived metabolite trimethylamine N-oxide is elevated in Alzheimer��s disease
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6303862/��
The Gut, the Heart, and TMAO
CLEVELAND HEARTLAB AUGUST 1, 2016 HEART ATTACK AND STROKE, TMAO
Dietary Choline-Gut Microbiome,The Surprising Link Between the Gut and Heart Health
Science has long recognized that what we eat plays a critical role in our heart health. Now the details of this complex connection are coming into focus.
One of the more intriguing recent discoveries has to do with the role of the gut microbiome��the trillions of microbes that reside in the GI tract and influence health by helping digest food, making vitamins, and providing protection against disease-causing microorganisms. Recently Cleveland Clinic researchers reported findings from several studies involving people and animals, that the gut microbiome directly changes the function of blood platelets, influencing the risk for heart attack and stroke.
Here��s how it works: When people ingest certain nutrients, such as choline (abundant in red meat, egg yolks, and dairy products) and L-carnitine (found in red meat as well as some energy drinks and supplements), the gut bacteria that break it down produce a compound called trimethylamine (TMA). The liver then converts TMA into the compound, trimethylene N-oxide (TMAO).
The trouble with TMAO is that data show high levels contribute to a heightened risk for clot-related events such as heart attack and stroke��even after researchers take into account the presence of conventional risk factors and markers of inflammation that might skew the results. In their most recent analysis, scientists showed that high blood levels of TMAO were associated with higher rates of premature death in a group of 2235 patients with stable coronary artery disease. Those found to have higher blood levels of TMAO had a four-fold greater risk of dying from any cause over the subsequent five years.
The implications are intriguing. Taken together, the new studies suggest that positively altering the gut microbiota may help to reduce damage to blood vessels, resulting in a stronger cardiovascular system, and they point to targets for potential new heart disease therapies.
The insights also underscore the potential power of TMAO testing, which can help patients determine if their gut microbiome is contributing to their risk for heart disease and whether they might benefit from limiting foods that contain the building blocks of TMAO. TMAO tests are only available through the Cleveland Heartlab.
To lower your TMAO levels, consider minimizing the consumption of full-fat dairy products, including whole milk, egg yolk, cream cheese, and butter; both processed and unprocessed red meat (beef, pork, lamb, and veal), as well as nutritional supplements and energy drinks containing choline, phosphatidylcholine (lecithin), and/or L-carnitine. Vegetarians and vegans, who avoid meat products, for instance, produce little TMAO.
In general, consuming a diverse diet rich in plant foods and fiber may be helpful. When Cleveland Clinic researchers fed mice a diet rich in TMAO producing nutrients, they identified a compound called DMB capable of minimizing TMAO produced from their gut microbiota. In fact, when DMB was added to their drinking water, they found TMAO levels and the formation of arterial plaques both declined. DMB may be found naturally in many Mediterranean diet foods, including red wine and extra virgin olive oil.
Such an eating pattern may turn out to be key to cultivating a healthy human gut microbiome��one that will fend off myriad illnesses, including heart disease, America��s chief health threat.The Gut, the Heart, and TMAO - Cleveland HeartLab, Inc.
https://www.clevelandheartlab.com/blog/the-gut-the-heart-and-tmao/��
Could Food Be a New Medicine to Fight Heart Disease?
CLEVELAND HEARTLAB JANUARY 11, 2016 DIET, HEART ATTACK AND STROKE, METABOLIC SYNDROME
A compound called DMB (3,3-dimethyl-1-butanol), found in olive oil, red wine and other foods, may someday be a first-of-its kind drug with the potential to treat��or even prevent��heart disease in the future, suggests a new Cleveland Clinic study published in the journal Cell.
The investigators report that in mice, dietary supplementation with this naturally occurring compound safely inhibited atherosclerosis (plaque buildup in arteries) and significantly lowered production of TMAO (trimethylamine-N-oxide), a heart-harming metabolite derived from gut bacteria.
If future studies show similar effects in people, the research may ultimately lead to a new strategy to treat��or even prevent��heart disease and stroke, two of the leading causes of death globally. To date, no trials of DMB have been done in humans.
��This study shows for the first time that one can target a gut microbial pathway to inhibit atherosclerosis,�� says senior study author Stanley Hazen, MD of the Cleveland Clinic. ��This new approach opens the door to the concept of drugging the microbiome to affect a therapeutic benefit in the host.��Could Food Be a New Medicine to Fight Heart Disease? - Cleveland HeartLab, Inc.
https://www.clevelandheartlab.com/blog/horizons-could-food-be-a-new-medicine-to-fight-heart-disease/��
3,3-Dimethyl-1-butanol (DMB) is a structural analog of choline.[1]
Effects
DMB inhibits microbial trimethylamine (TMA) formation in mice and in human feces, thereby reducing plasma trimethylamine N-oxide (TMAO) levels after choline or carnitine supplementation.[1] It consequently inhibited choline-enhanced endogenous macrophage foam cell formation and atherosclerotic lesion development in mice without alterations in circulating cholesterol levels.[1]
While mice placed on a choline supplemented diet showed an increase in the proportions of the bacterial taxon Clostridiales in the gut, DMB induced a decrease in the proportions of this taxon.[1]
Mice showed no evidence of toxicity to chronic (16-week) DMB exposure.[1]
3,3-����-1-������DMB���ǵ���Ľṹ�����[1] ��Ч DMB����С������������������װ���TMA�����γɣ��Ӷ����Ͳ��䵨�������Ѫ�����װ�N-�����TMAO����ˮƽ��[1]��ˣ���������С����ǿ����Դ�Ծ���ϸ����ĭϸ�����γɺͶ�������Ӳ������ķ�չ�������̴�ˮƽȴû�иı䡣[1] ���ܵ������ʳ��С����ϸ������Ⱥ��״ѿ���˾��ı������ӣ���DMB�յ��÷���Ⱥ�������͡�[1] С��δ��ʾ�����ԣ�16�ܣ�DMB��¶�ж��Ե�֤�ݡ�[1]3,3-Dimethyl-1-butanol (choline analogue) as a TMAO inhibitor - Supplements - LONGECITY
https://www.longecity.org/forum/topic/103257-33-dimethyl-1-butanol-choline-analogue-as-a-tmao-inhibitor/��
EPA and DHA for IBD
4. Inflammatory Bowel Disease (IBD)
The same or similar story is being described in the research literature last year or this year for essentially all of the inflammation-related diseases. The 2016 publication Inflammatory bowel disease: can omega-3 fatty acids really help? relates: ��Adjuvants to the traditional therapy of inflammatory bowel disease (IBD) have been studied to enhance the efficacy of the treatment and improve patients�� quality of life. Omega-3 polyunsaturated fatty acids (��3FA) have been associated with attenuation of the inflammatory responses in IBD, possibly acting as substrates for anti-inflammatory eicosanoid production, similar to prostaglandins and leukotrienes. ��3FA also act as substrates for the synthesis of resolvins, maresins and protectins, indispensable in resolving inflammation processes. These acids may influence the development or course of IBD by: reducing oxidative stress, production of tumor necrosis factor-�� and proinflammatory cytokines; working as chemopreventive agents; and decreasing the expression of adhesion molecules. There are numerous controversies in the literature on the effects of ��3FA in the prevention or treatment of IBD, but their effects in reducing inflammation is incontestable. Therefore, more studies are warranted to elucidate the pathophysiological mechanisms and establish the recommended daily intake to prevent or induce remission in IBD patients.��
This 2016 publication relates to inflammatory bowel disease, EPA- and DHA-derived resolvins�� actions in inflammatory bowel disease. ��Inflammatory bowel diseases are chronic diseases divided into two major forms, ulcerative colitis and Crohn��s disease, which are both associated with a chronic inflammatory condition of the gastrointestinal tract. Recent studies have shown that the resolution of inflammatory conditions is a biosynthetically active process where new pro-resolution lipid mediators derived from omega-3 fatty acids eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA), such as E- and D-series resolvins, protectins, and macrophage mediator in resolving inflammation (maresins), have potent anti-inflammatory activity and serve as specialised mediators that play an important role in the resolution of inflammation. Recent studies have also shown the role of resolvins in referred hyperalgesia associated with different inflammatory processes, such as the visceral pain caused by inflammatory bowel disease. There are many reports describing the principal effects of EPA- and DHA-derived mediators in experimental models of inflammatory bowel diseases. This review focuses on the recent studies on the important role played by pro-resolution lipid mediators in controlling the inflammatory process associated with inflammatory bowel diseases.��
��
Types of Diarrhea
There are 3 basic categories of diarrhea: watery, fatty (malabsorptive) and inflammatory. Not all diarrhea is purely watery, malabsorptive, or inflammatory because some categories overlap. Also, individuals may experience diarrhea which has multiple causes, thus presenting a more complex situation.
1. Watery Diarrhea
Watery diarrhea is characterized by runny or watery bowel movements. There are 3 subcategories of watery diarrhea: osmotic, secretory and functional.
Osmotic Diarrhea
Osmotic diarrhea occurs when something is drawing water from the rest of the body into the bowels. In most cases, osmotic diarrhea will cease when one stops ingesting the food or substance that is causing water to be drawn into the bowels.
Secretory Diarrhea
Secretory diarrhea involves a higher stool volume, diarrhea that often occurs through the night and will continue even if one in fasting.
Secretory diarrhea occurs when more water is released into the bowels than is usual. This happens due to an increase in secretion or a decrease in absorption of electrolytes in the bowels. There are a variety of causes for this disruption.
Functional Diarrhea
Functional diarrhea involves hypermotility (excessive movement of the muscles in the GI tract) and is characterized by smaller volumes of diarrhea. Functional diarrhea improves at night and with fasting. Functional diarrhea does not have any associated abdominal pain.
People with functional diarrhea don��t usually have it all the time. Normal bowel movements, and even constipation can occur between bouts of diarrhea.
Functional Diarrhea has no know cause. There are no structural or biochemical abnormalities with this type of diarrhea, part of the GI system just does not function quite right.
2. Fatty or Malabsorptive Diarrhea
Malabsorptive diarrhea occurs when the digestion or absorption of nutrients is impaired. It often includes the presence of excess fat in the stool (appearing as frothy or greasy stool), a foul odor to the stool, and symptoms such as bloating, gas, and weight loss.
Malabsorptive diarrhea improves with fasting.
3. Inflammatory or Exudative Diarrhea
This type of diarrhea results from a damaged intestinal lining. Inflammation is usually present in the bowels, which is why it is described as inflammatory. There is often blood or pus present in the stool so it is sometimes referred to as exudative diarrhea.��
�������
��к�����ֻ�������:ˮ����к��֬���Ը�к(���ղ���)����֢�Ը�к�����������еĸ�к����ˮ���ģ����ղ����ģ���������֢�Եģ���Ϊ��Щ�������ص��ġ����⣬������ܾ�����к�Ķ���ԭ����˳��ָ����ӵ������
1. ˮ����к
ˮ����к�������dz�����ˮ���˶���ˮ����к�ɷ�Ϊ���ԡ������Ժ����������ࡣ
���Ը�к
��ijЩ���������������������ȡˮ�ֽ��볦��ʱ���ͻᷢ�����Ը�к���ڴ��������£���һ����ֹͣ���뵼��ˮ���볦����ʳ�������ʱ�����Ը�к�ͻ�ֹͣ��
�����Ը�к
�����Ը�к������������ӣ���кͨ����������������ʹ��ʳҲ�������
�����Ը�к����ʱ�������ˮ���ͷŵ�������ƽʱ�������������ӷ��ڻ�������յĵ�����ڳ�����������ֻ��ҵ�ԭ���кܶࡣ
�����Ը�к
�����Ը�к����θ������������˶����Ը�к����СΪ�����������Ը�к��ҹ��ͽ�ʳʱ�������ơ������Ը�кû���κ���صĸ�ʹ��
���й����Ը�к����ͨ������һֱ������֢״�������ij����˶�����������Ҳ���ܷ����ڸ�к֮�䡣
�����Ը�к���������������͵ĸ�кû�нṹ�������쳣��ֻ�Dz���θ����ϵͳ���ܲ�̫������
2. ֬���Ի����ղ����ĸ�к
��Ӫ�����ʵ������������ܵ���ʱ���ͻᷢ�����ղ�����к����ͨ����������еĶ���֬��(����Ϊ��ĭ������ķ��)�����Ķ�����Լ����縹�͡����������ؼ����֢״��
��ʳ�ɸ������ղ�����к��
3.���Ի������Ը�к
�������͵ĸ�к���ɳ����ڱ���������ġ���֢ͨ�������ڳ����������Ϊʲô������Ϊ��֢������о�����Ѫ��ŧ��������ʱ����Ϊ�����Ը�к����
Digestive System Disorders:
Dietary Intolerance �C Conditions such as celiac disease (gluten intolerance), lactose intolerance, and fructose intolerance are caused by enzyme deficiencies or abnormalities in the intestinal mucosa, this impairs the necessary functions of the GI tract and leads to malabsorption of nutrients. When nutrients are not absorbed properly, they build up in the intestine and draw fluids to the intestines. Such conditions can cause secretory or malabsorptive diarrhea.
Bile Acid Deficiency �C A reduced amount of bile acids will impair the absorption of fats resulting in fatty or malabsorptive diarrhea. Low bile acids can be due to a number of causes including decreased liver or gall bladder function, low cortisol levels, or prescription medications. Bile acid deficiency is different than bile acid malabsorption, which involves excess secretion of bile acids into the bowels, resulting in secretory diarrhea.
Pancreatic Insufficiency �C The loss of pancreatic secretions which occurs in cystic fibrosis and pancreatitis also effect digestion and absorption of nutrients.
Irritable Bowel Syndrome (IBS) �C Functional diarrhea is one of the symptoms experienced by people with IBS. Functional diarrhea and IBS are often mistaken for the same condition, but they are not since IBS includes other symptoms, such as abdominal pain.
Inflammatory Bowel Diseases �C There is usually a lot of damage to the intestinal wall in inflammatory conditions such as Chron��s disease, ulcerative colitis, and diverticulitis. Exudative diarrhea is most common in such conditions. Secretory diarrhea occurs in microscopic colitis and in earlier stages of some of the other inflammatory bowel diseases.
����ϵͳ���µĸ�к:
��ʳ����֢����������к(�ȵ��ײ���֢)�����Dz���֢���Dz���֢�����ɳ���ճĤ��øȱ�����쳣����ģ�������θ�����ı�Ҫ���ܣ�����Ӫ�����ʵ����ղ�������Ӫ������û�б���ȷ������ʱ�����Ǿͻ��ڳ����жѻ�������Һ�����볦������������ᵼ�·����Ի����ղ����ĸ�к��
��֭���-��֭��ļ��ٻ���֬�������գ�����֬���Ի����ղ����ĸ�к����֭��͵�ԭ���кܶ࣬����������ҹ����½���Ƥ�ʴ�ˮƽ�ͣ���ҩ����֭��ȱ����ͬ�ڵ�֭�����ղ����������ǵ�֭����ڹ�����볦�������·����Ը�к��
���ٹ��ܲ�ȫ-������ά�����������µ����ٷ��ڼ���Ҳ��Ӱ��Ӫ�����ʵ����������ա�
�����ۺ���(IBS) -�����Ը�к��IBS���߾�����֢״֮һ�������Ը�к�ͳ����ۺ�֢����������Ϊ��ͬһ�ּ�������ʵ���ϲ�����ˣ���Ϊ�����ۺ�֢����������֢״���縹ʹ��
��֢�Գ�����������֢״̬�£������Լ����������Խ᳦�������ף�ͨ����Գ�����ɺܴ�������������£������Ը�к����������Ը�к�����������½᳦��һЩ������֢�Գ��������ڽΡ�https://flowingfree.org/diarrhea-no-more-leave-the-toilet-behind/
https://www.ibstales.com/caltrate-plus.htm
Ingredients in Caltrate plus
��
��
����ҽѧԺ �����ϸ���о�Ժ 2017
�˽⳦��������滻����������
�����ѧ���о���Ա���֣���С���У�����ϸ�����Իָ�Ϊ��ϸ�����Դٽ���֯��������
�����ϸ���о���������-������֢�о�������ҽѧԺ���о���Ա������һ��֮ǰδ֪�Ļ��ƣ����ڳ����ڲ������з�����Ҫ���á����ǵķ���Ϊ���ÿ�춼�ڷ����仯����֯���ά�������ṩ���µļ��⡣
С��������������������ǿ�����٣�ÿ5��7������һ�ν�����Ƥ����Ĥ��ϸ���ij�������ʹ��Ƥϸ���ܹ������ڷֽ�ʳ�����Ӫ������������Ĺ����������ܵij���ĥ��
��һ��ϸ����ϸ���о��У��о���Ա���ֳ���ϸ����������������ϸ��������С�������и�ϸ����������
������һ��dz������ķ��֣����������ܹ������˽���֯�������֯�����ģ��������ϸ���о���(Harvard Stem Cell Institute)����ϯ��Ա������-������֢�о���(Dana-Farber Cancer Institute)����ҽѧԺ(Harvard Medical School)��ҽѧ���ڡ����о�������������÷ʲ��ϣ�������(Ramesh Shivdasani)˵��
�ڹ�ȥ��10�����ѧ����һֱ��Ϊ��������Ƥϸ�������ij̶�ȡ������һ����ϸ���Ĵ��ڣ���Щ��ϸ������Ҫ��ʱ����Ȼ��������״̬���෴�������������о�����������ϸ�����ľ�ʱ��һЩ���͵ij���ϸ���ھ�����һ������ȥ�ֻ��Ĺ��̺�Ὣ�Լ�ת��Ϊ��ϸ����
����������ǿ��ܵģ���Ϊ�����������е�ϸ��������ͬ���Ŵ����룬��Shivdasani˵��������ʹÿ��ϸ����ͬ������Щʵ�ʻ�Ծ���Ŵ�����IJ��֡���
Ҫ�뷢��ȥ�ֻ���De-differentiation)��ϸ����Ҫ����������DNA��ϸ�������۵���Ⱦɫ�ʵķ�ʽ���⽫�ı���Щ�����ǻ�Ծ�ģ���ͨ������Ϊ�Dz����ܷ����ġ�
Ȼ�����ںĽ���С�������е�ԭʼ��ϸ����Shivdasani������ͬ���Ƿ�����ÿ��ϸ���������еķ��ӱ�Ǻ�Ⱦɫ��ǩ������ȷ��������Щ���͵�ϸ����������
���Ƿ��֣������¸�ϸ���ij��֣����ֲ�ͬ���͵�ϸ��Ⱥ�����˱仯���������Щϸ������ȥ�ֻ�����Ϊ��ϸ��������֤�ݶ�������ͬ���Ļ���Ҳ���ܷ������������ϡ�
�������ƺ����о�Ŀ����ԣ���Shivdasani˵����������ǧ�Ƶij������У����ǿ��Կ���Ⱦɫ�ʵ�չ������
�����о���ʾ�˷�����������֯��Ļ������̣�ʹ���ǵ������ܹ��������ˣ�������Ϊδ��ȷ�����ưе��ṩ������Ȼ����Shivdasaniָ����Ҫ�˽���Щ֪ʶ������������ض����������ƻ�Ϊʱ���硣
��������Ȼ�����׳��ڵij���ϸ�������֪����ϸ��ȱʧ�ģ���Shivdasani˵�����������Ѿ�������Ⱦɫ��������������ģ����ڽ���ȥ�ֻ���ϸ�����Ա�������о�������һ����Ҫ�Ŀ�ʼ����
�����о��õ��˹����������������༲���о��������ҹ�����Ⱦ�����о��������������о������������Լ����ڷ��������о������ͷ�������ս��֯�ľ�������
��̾~����������������� ��ҽ "ƢθΪ����֮������Զ����
����~��Ȼ����ij�����Ƥϸ�����ԡ�ȥ�ֻ�������Ϊ��ϸ��������ϸ���İ������ͨ��ϸ������Ϊ����֢��ϸ������Ҳ�Ͳ��������ˡ�
Understanding how the intestine replaces and repairs itself �C Harvard Gazette
https://news.harvard.edu/gazette/story/2017/07/understanding-how-the-intestine-replaces-and-repairs-itself/��
Autism and the Human Gut Flora
Previous research into autism has raised the possibility that a link exists between the disorder and the human gut microflora. Research has remained limited due to the unwillingness of the orthodox medical establishment to adopt treatments suggested by this research. This short review aims to summarise the research completed to date and evaluate the relevance of a link between autism and a possible imbalance in the human gut microflora. It appears that yeasts (Candida species in particular) and clostridia may play an important role in the development of autistic symptoms. It is suggested that the control of the growth of these species may reduce the severity of autistic symptoms but is unlikely to offer a cure.
Introduction
Previous research into autism has raised the possibility that a link exists between the disorder and the human gut microflora. However the relevance of this to sufferers of Autism has remained somewhat un-researched. As a consequence, it appears that the treatments suggested previously have remained unused and regarded by the orthodox medical establishment as irrelevant. This short review will consider the limited research completed to date.
Autism
The term autism is usually associated with the syndrome first described by Kanner (1943). In more recent years specific criteria have been set out to aid in the diagnosis of the disorder (Shaw et al, 1999). Autism typically develops early in childhood, however causality, explanation and treatment are often hotly debated. Symptoms can include (but not necessarily), hyperactivity, loss of eye contact, decreased vocalisation (i.e. loss of language), stereotypical behaviours, poor academic performance and other similar social deficits. Other similar disorders exist. These include Asperger Syndrome, Attention Deficit Hyperactivity Disorder (ADHD), Pervasive developmental disorder (PDD) and many others, where symptoms are similar to Autism but specific differences are demonstrated.
Yeast metabolites in the Urine of Autistic Children
Yeasts constitute only a very small proportion of the population of the gut (Holzapfel et al, 1998) under normal conditions �C possibly kept from growing via competition from bacterial species and certain immune functions. However, Shaw et al (1999) has proposed that Autism (or at least many of the symptoms) may be a consequence of an overgrowth of candida species and a selective IgA deficiency. Following treatment with antifungal drugs and a gluten and casein free diet, a child rated as having severe autism improved such an amount as to be classed as a higher functioning individual with autism.*
It has been shown previously that children exhibiting autistic features have increased excretion of arabinose and the analogs of Krebs Cycle metabolites (including tartaric acid) (Shaw, Kassen & Chaves, 1995). Using Gas Chromatography/Mass Spectrometry (GC/MS) to test for urinary metabolites, it was found that the children had extremely high values of tartaric acid. The only source of tartaric acid is yeast (Shaw, 1999). Many reports have suggested that the onset of autism may be related to the occurrence in children of otitis media (Kontstantareas & Homatidis, 1987). It is common to treat otitis media with some sort of broad-spectrum antibiotic. Intestinal overgrowth of yeast and certain anaerobic bacteria are a well documented outcome of the administration of broad spectrum antibiotics (Kennedy & Volz, 1983; Danna et al, 1991; Ostfield et al, 1977; Kinsman et al, 1989; Van der Waaij, 1987; Samsonis et al, 1993, 1994a,b).
To evaluate this, it is known that Shaw (1999, 1996) has used GC/MS to test the urinary content of metabolites following the administration of Nystatin, an antifungal drug. Accordingly it was found that urinary tartaric acid declined to zero after about 60 days. When the Nystatin dose was cut to half, levels appeared to rise. Associated with this was seen an improvement in eye contact, a reduction in hyperactivity and an improvement in sleep patterns. It is unclear as to the method used, however these are interesting results. In other work, Shaw (1999) has evaluated the progress of the Herxheimer reaction of yeast die off. Values for the microbial metabolites described above increased dramatically during the first three days and began to normalise near day four. It is unclear as to the relevance of the point that many children take antibiotics for a whole series of illnesses, but do not develop autism.
Gupta et al (1996) have studied some possible modulating factors in Autism. These include immunodeficiencies and probable differences in biochemical detoxication factors which appear to be very common in autism,. It has been estimated that a high percentage of autisitic children have a significant immune dysfunction that may include myeloperoxidase deficiency, a genetic deficiency that impairs the action of white blood cells on yeast cells, IgA deficiency, complement C4b deficiency, IgG deficiency or IgG subclass deficiency (Gupta et al, 1996). In the above study, a complete remission of autisitic symptoms was achieved by infusions of gamma globulin in one case. Shaw (1999) has suggested that it seems increasingly likely that the immune system takes an inventory of bacteria and yeast cells present in the gut soon after birth. This inventory is performed by CD5+ B-cells. These cells may play a role in regulating the secretion of IgA, the antibody class that is secreted into the intestinal tract and which may select which microorganisms are tolerated in the gut. Continuing, it is suggested that the eradication of the normal gut flora when antibiotics are administered repeatedly during infancy may cause the CD5+ cells to reject normal cells and award immune tolerance to species that potentially could cause harm. Either antibiotic use in infancy or yeast infection of the mother during pregnancy may result in later tolerance to yeast. This would possibly extend any remission from symptoms induced by methods of treatment suggested for autism.
Yeast and Tartaric Acid Toxicity
Shaw, Kassen & Chaves (1995) noted in their initial study of the two brothers with autism, that in addition to their autistic characteristics, they exhibited severe muscle weakness. Both of these brothers excreted large amounts of tartaric acid in their urine. It is known that tartaric acid is an analog of malic acid. Malic acid is a key intermediate in the Krebs cycle which is responsible for the extraction of most of the energy from food substrates. Tartaric acid is presumably seen as toxic since it would inhibit this pathway and limit energy production. This may be the reason for the muscle weakness seen in the two autistic brothers. It has also been shown that Candida albicans produce gliotoxins (Shah and Larsen, 1991 and 1992) and immunotoxins (Podzorski et al, 1989; Witken, 1985), which may further impair the immune system. This would have relevance in terms of promoting yeast growth and increasing the chances of additional infections from bacteria leading to antibiotic usage again.
Arabinose, Yeast and Autism
Shaw, Kassen & Chaves (1995), also identified high levels of arabinose in the urine of the autistic children. The exact biochemical role of arabinose is unknown, but a closely related yeast alcohol, arabitol has been used as a biochemical indicator of invasive candidiasis (Kiehn et al, 1979; Wong et al, 1990, Roboz & Katz, 1992). It is thought that arabitol produced by the yeast in the gut is absorbed into the portal circulation and then converted to arabinose by the liver. Elevated protein-bound arabinose has been found in the serum glycoproteins of schizophrenics (Varma & Hoshino, 1980) and in children with conduct disorders (Varma et al, 1983).
Arabinose reacts with the epsilon amino group of lysine in a wide variety of proteins and may then form cross-links with arginine residues in an adjoining protein (Sell & Monnier, 1989). Arabinose has therefore been implicated in protein modification via cross-linking the proteins and altering both biological structures and functions of a wide variety of proteins. Shaw et al (1999) has proposed that this may include proteins involved in the interconnection of neurons. Decreased clinical symptoms of autism after antifungal treatment might be due to decreased arabinose and pentosidine formation (cross-linked proteins described above). This would result in fewer random neural connections and increased numbers of neural connections that are oriented to the child��s environment. The influence of a number of vitamins on candida activity and the operation of many metabolites of candida has been discussed (Mahler & Cordes, 1966). It is commonly found that children with autism experience improved symptoms following removal of gluten and casein from the diet (Shattock et al 1990, 1991, 1996, 1997; Reichelt et al, 1981, 1986; Knivsberg et al, 1990). It is unclear whether there is a link with arabinose and pentosidine formation, however given the wide distribution of these proteins in foods, there may be a relevant process involved with respect to autistic symptoms. Research into this may be useful.
Clostridia and Autism
Bolte (1998) outlined the possibility of a subacute, chronic tetanus infection of the gut as the underlying cause for symptoms of autism observed in some individuals. Here it is postulated that a percentage of individuals with autism have a history of extensive antibiotic use. As above it is known that oral antibiotics significantly disrupt protective gut microflora, creating a favourable environment for colonisation by opportunistic pathogens. Clostridium tetani is a ubiquitous anaerobic bacillus that is known to produce a potent neurotoxin. The normal site of binding for tetanus neurotoxin is the spinal cord, yet the vagus nerve is capable of transporting tetanus neurotoxin, providing a route of ascent from the intestinal tract to the central nervous system and bypassing the spinal cord. The result would be the typical symptoms of a tetanus infection would not be evident. Once in the brain the tetanus neurotoxin disrupts the release of neurotransmitters. This may explain the wide variety of behavioural deficits apparent in autism. Bolte (1998) presents evidence of lab animals exhibiting many of these behaviours after being injected in the brain with tetanus neurotoxin, and also of children with autism showing a significant reduction in sterotyped behaviour following treatment with antimicrobials effective against intestinal clostridia.
Shaw (1999) has developed a similar technique to yeast metabolite analysis by GC/MS for clostridia metabolites. While it has not been fully identified it has been shown that tyrosine derivative, a compound very similar but not identical to 3,4-dihydroyphenylpropionic acid is raised in the urine of children with various conduct disorders. It has also been pointed out that tyrosine, a substrate used by the body for the production of neurotransmitters, might imply that this unidentified product is important in altering key biochemical pathways for neurotransmitters in the brain. The relevance to autism remains unclear. Clearly a treatment to reduce the numbers of clostridia in the gut might help to reduce the effects of this inhabitation. However, clostridia are spore forming meaning recolonisation is possible even after treatment. The relevance of probiotic and prebiotic treatments has not been researched, yet this appears a reasonable approach to aid suppression of the clostridia species.
Human Gut Flora and Gluten and Casein Intolerance in Autistic Children
Yeasts, including Candida albicans are known to secrete a number of enzymes. These may include phospholipase, which will break down phopholipids and proteases such as secretory aspartate protease which break down proteins. These enzymes may partially digest the gut membranes and lining itself. Furthermore, it is known that the mycelium and chlamydospore are capable of tissue invasion (Nolting et al, 1994). It is likely that such factors could increase the permeability of the gut. This has important consequences in terms of food absorption and digestion. In terms of autism symptoms this may be highly relevant. It has been shown that incompletely broken down portions of gluten and casein may be crossing the gut into the blood and having an opioid effect in autistic children. Their symptoms being a consequence of this opioid action (Reichelt, 1981; Shattock et al, 1990). While many mechanisms have been suggested for this incomplete breakdown, it seems key that a yeast and/or clostridia overgrowth would affect this in some way.
Conclusions
Products of the human gut microflora in relation to autism and its symptoms appear to have been largely ignored in the past. However they appear to be relevant certainly in terms of yeast and clostridia species. Abnormal bacterial metabolites of tyrosine appear to be elevated in autism and this seems related largely to an overgrowth in clostridia species. Treatments to control this have resulted in significant clinical improvement or complete remission of symptoms in some cases. The issue of whether probiotic and prebiotic treatments are relevant, needs to be researched. Elevation of yeast metabolites such as tartaric acid and arabinose appears to be even more common in autism. The arabinose appears to be involved in abnormal protein binding which may adversely affect neuron connection and this may have relevance to the appearance of autistic symptoms. Given the intolerance autistic children appear to have to gluten and casein, an involvement of arabinose may be important. However, the exact biochemical role of arabinose remains unclear. Tartaric acid from the yeast overgrowth may have a direct toxic effect on the muscles and is a key inhibitor of the Krebs Cycle that supplies raw materials for gluconeogenesis. The implications of this appear to be detrimental to autistic function.
While it is clear that abnormal metabolites from the human gut flora may contribute to autistic symptoms, it is certain that many other systems and pathways are involved. It is possible that many are functioning simultaneously but at varying levels between individuals. This may give rise to the differences in symptoms exhibited by sufferers. This may also explain the similarity of autism and other chronic behavioural disorders such as Asperger Syndrome, PDD, ADHD, ADD and Rett syndrome. While other pathways may exist, research into the role of human gut flora and the full identification of species involved may allow for more directed treatments. While it will not cure the disorder, modifications of gut flora function might improve symptoms significantly.
* The Childhood Autism Rating Scale, an observational measure of various aspects of autism, for the child in question decreased from 43 (severely autistic) prior to introduction of these therapies to a value of 29 (non-autistic) after therapy.Autism and the Human Gut Flora
https://www.ei-resource.org/articles/autism-articles/autism-and-the-human-gut-flora/?tmpl=component��
��
.gif)

.png)

.png)