��
ά����C�;���ϸ����GcMAF
��
ά����C���ӻ����ϸ����һ���������γ�
Ascorbate-Dependent Enhancement of Nitric Oxide Formation
in Activated Macrophages
ժҪ
��ϸ��֬����(LPS)��ϸ�����Ӽ���ľ���ϸ��ͨ���յ�iNOS����������һ��������NO)���ڸ�����-�ú�LPS������������ϸ����ϸ��ϵJ774.1,����Ѫ�������ӵ�������(�������κ�������)���γɴ�Լ40%������ʹ�ÿ���Ѫ���NO���γ����յ����á�
NGһ���������������л�����Ѫ�ἤ���ϸ�������������ε��γɡ�
Western blotting������ʾ���ڿ���Ѫ�ἤ���ϸ���У�iNOSmRNA�͵����ʵ���̬ˮƽ���Ƚ����յ��D���ϸ���������˴�Լ������
��Щ���ݱ���������Ѫ��ͨ�����ӻ����ϸ����iNOS������������NO�IJ�����
��
��
Ascorbate-Dependent Enhancement of Nitric Oxide Formation in Activated Macrophages��
Abstract Macrophages activated with bacterial lipopolysaccharide (LPS) and
cytokines produce nitric oxide through the induction of iNOS gene expression.
Ascorbate increased NOx (nitrite and nitrate) formation by approximately 40% in
a mouse macrophage-like cell line, J774.1, activated with LPS and interferon-��.
Ascorbate alone exhibited no inductive activity toward NO
formation.NG-Monomethyl-l-arginine inhibited nitrite formation in cells
activated in the presence or absence of ascorbate. Northern and Western blotting
analyses showed that both iNOSmRNA and protein steady-state levels were
increased approximately twofold in cells activated in the presence of ascorbate
compared to in cells activated only with the inducers. These data suggest that
ascorbate increased NO production by increasing the amount of iNOS in the
activated macrophages.
��
Ascorbate-Dependent Enhancement of Nitric Oxide Formation in Activated
Macrophages��
https://doi.org/10.1006/niox.1998.0182
��
NO-dependent biological actions of human macrophages

��
����ϸ������һ������:���Ǻ������ʵ�
Nitric oxide production by human macrophages: there's NO doubt about it

NO�ϳ�ø��l-������������һ������(NO)�ķ��ֳ��ı������Ƕ���������ѧ���������⡣�Դ�Furchgott��Zawadzki(7)�Ŀ������о�������NO������NOѭ�����õ�����ҩ��ѧ�Ƽ������ٴ��õ��㷺Ӧ�á�Ȼ��������NO�ڿ���Ⱦ�е������γ��˶Աȣ����ߵ��ٴ�Ӧ���ںܴ�̶��ϻ�û�б���ʶ����Hibbs���˵Ŀ������о�(8)֤����С�����ϸ���ķ����������ã���һ�о��ѱ�֤����������ƶϸ�Ϊ���ѡ�
������֤������ϸ���ܷ����NO�����������ϰ����������ʣ��ڸ�Ⱦ��������֢����£�����NO�ķ�������������(4,24)��Ȼ��,�������������Ѫ����ϸ����֬����interferon-��δ������û������(15),��ʹ��Щ�̼��Ǹ߶���Ч������С�����ϸ������������NO(19)�����һЩ�о��������������ϸ���ܹ�NO(15)��
�������Ĵ��ڹ�ȥ10���100�����о�������ˮ�档Ŀǰ���������ϸ������NO�Dz�������ʵ���ڸ��ֻ�����(10,14,25)���ڻ���˾���ϸ�����ظ�������NO��ømRNA�͵��ף�ͬʱҲ������֤�ݱ���NO�IJ���(23)���յ���һ��������ø(iNOS)�������ڸ�Ⱦ���߽�����֯����ϸ���еõ��������ŷ���֤��(1)��������о����֣�iNOS�����Ӷ�̬�������ϸ��NO�IJ����ͶԸ�Ⱦ�ĵֿ��й�(11)�����һ��֧����NO���������������е���Ҫ���á����˾���ϸ������ʾNO�Ĺؼ��������̼������ڷ�������Ȼ�ڻ�����֢�Ļ���(��ν��(12)�����ʪ�ؽ���(18)��ű��(2))�ľ���ϸ���о�������NO�����ڽ�������������Ѫ����ϸ���̼�NO�����������������Բ���ȫȷ����
����һ��(�ο�����9���μ���L944ҳ)�У�Hickman-Davis���˶Դٽ��������ϸ������NO�������б���������Ҫ���䡣һЩ��������ֲ���ߵķ��ݾ���ϸ���ڳ������½���֧���ܷ��ݹ�ϴ��������־Ը�ߵľ���ϸ��δ�����Է�����������NO���ڱ�����Ե���A
(SP-A)������ײ���(liveKlebsiella pneumoniae)�Ĵ̼��£���Щ����ϸ����һ����ǿ��ֵ��ע����ǣ���sp -
a������ϸ���Կ��ײ��Ͼ����п������ԣ�������NO�IJ��������������ֿ������ԡ����������ֲ�ķ�Ϊ���������ṩ��һ���ٽ��ֿ�ϸ����Ⱦ�Ĵ̼�����������һ��֤��������SP-A�����ļ���������Զ��ַ�ʽ�ٽ��������ߡ�����ֲ�оֲ�NO�ϳɵ�֤ʵ֤ʵ�����ڵ��о��������ͬ���������ֲ���ߵ�֧���ܷ��ݹ�ϴҺ�к���iNOS
mRNA(16)����Ȼϸ���Է����ڷ���ֲ����ںܳ���(3)�������ݾ���ϸ���ļ���������������ƴ˺���ķ�����
Hickman-Davis et al
.(9)�ij��Բ��������źŴ�ijЩǰ����ݾ���ϸ����ֲ���ߺϳ�û�л�ӦSP-A,���collectin��ǿ����ϸ�����ܺ����������û�������������ϸ���Ŀ������Բ�ȷ�䡣�����ײ��Ͼ������NO������SP-A�൱��˵��NO��SP-A�Ĺ������أ���������˵��SP-A�Ŀ������á����Ƶأ�sp
-
a�յ���ϸ����Ca2+�������ƺ�����Щ���õı�Ҫ������������ֲ���ߺ�����������ľ���ϸ���о��з�����ϣ����-��ά˹������ͬ��֤�����ɻ�����/����������øϵͳ������һ�������ͻ���������������Ѹ��ɱ�����ײ�������Ȼ�����ⲻ����ܽ������ڷ��ݾ���ϸ���й۲쵽����������Ҫ���־����ã������о���ԱҲû���ṩ����ϸ��������������֤�ݡ�����ϸ����Դ��NO���־�����ʵ������С��ϸ���й۲쵽��NO�����Կ�������һ��(22)��NOͨ�������������ͽ�������������Ʊ�Ҫ��ϸ�����̵�����Ϊ�־��ṩ��һ����еԭ��(17)������ֱ�ӵĿ��������⣬������Ϊ���ݾ���ϸ����NO����������NO�Ķ�Ч�Ի�����������й�(5,17)��֧����һ��˵���ǣ�NO��С������ײ��Ͼ���Ⱦģ���еı�������������Ϊ�����ʱ���ĵ��ڼ����������(20)��
NO������ֵ��غ�ϸ���������õ�����(6)���������ڸ�������ϵͳ(����ֲ��)�еľ�ȷ���ӻ��ƴ����˳�������ս��Ȼ����Hickman-Davis����(9)�Ĺ۲�ǿ����NO�������������ߵ���Ҫ�н����Ϣ��δ���Ĵ�Ⱦ�����Ʋ��Կ������Ϊ���Ӹ�Ⱦ��λNO�IJ������������к��ĸ�����(21)�������о���Ա����һЩʱ�䣬������ϸ������ѧ���˾ܾ���
Nitric oxide production by human macrophages: there's NO doubt about it |
American Journal of Physiology-Lung Cellular and Molecular Physiology
https://www.physiology.org/doi/full/10.1152/ajplung.00017.2002
��
һ�������ϳ��ھ���ϸ�����������е�����
��ȥ5����о�������С�����ϸ�����������յ���һ��������ø�Ļ��ԣ���ø����l
-��������ҵ��������ϰ���ͻ������ɻ�һ���������Ծ���ϸ���и�ͨ·�����ﻯѧ�����ߵ��ڵ�����Ϊ������ЧӦ�����ṩ�˱�������ȥ2�귢���ı��������һ�������ĺϳɽ鵼��С�����ϸ����ijЩ�������桢ԭ��������²����Ŀ������ԡ�
Role of nitric oxide synthesis in macrophage antimicrobial activity
Research over the past 5 years has demonstrated that immunologic activation of
mouse macrophages induces the activity of nitric oxide synthase, which oxidizes
a guanidino nitrogen of L-arginine, yielding citrulline and the reactive
radical, nitric oxide. A review of the biochemistry and immunologic regulation
of this pathway in macrophages provides a backdrop against which to evaluate its
effector functions. Reports published in the past 2 years suggest that synthesis
of NO mediates much of the antimicrobial activity of mouse macrophages against
some fungal, helminthic, protozoal and bacterial pathogens.
Role of nitric oxide synthesis in macrophage antimicrobial activity -
ScienceDirect
https://www.sciencedirect.com/science/article/abs/pii/095279159190079G
��
��
GcMAF���飬������
��ľ���ϸ��ɱ��
Your Awesome Macrophage Killing Machine
����ϸ���Ǿ�ġ������İ�ϸ�������𡢲������ɲ����������ߡ������ղ�������(������˼�ǡ��Ե���)���ǵĵ��ˡ����ǿ����ڱ�ҪʱѸ�ٷ�ֳ��Ȼ���������������裬��Ҫ��GcMAF���Opsonin��������ˮ����������ճס������ǵĵ������������ɻ���������(�ֳơ�������ը��)��������Ͱ�ϸ����ը������һ��һ�������ϸ�����������ɣ����ͱ���װ��һ����������ø�塱(ϸ���ڵġ�����ǻ��)�У�����������ɱ��(�������û�����Ļ�)��Ȼ�ֽ��������ɲ��֣�Ȼ��ѭ�����á�
��Ȼ���Ѿ������˾���ϸ��������Щ��������ϸ����������GcMAF��ι����dz���Ҫ���������Ҫ����ϸ�ؽ������ǡ����⣬�������������˵Ķ���!
�����������һ������,������,����,����ؿ��µľ��ޱȵ�ճ��blob��յ�Σ�յĶ�ħ���������ϵ���в��С,�����Lechterͬ����ʳ,��˹��ά��Ĺ⽣,�ս���,��С��,���ǵ���,��ŦԼGhostbusters-all����һ�����ɱ¾��������һЩ�뷨�ľ���ϸ�������Ǵ���������������ġ��������ϸ����������ϸ������ᾡ���Ŭ���ܿ����ǡ�
����ϸ���Ǿ��
���ް��ж��?���֮�£���ϸ������ϸ���͵��͵İ�ϸ��ֱ��ԼΪ7��(������֮һ��)�����ԼΪ250�����ס�����ϸ��ֱ��ԼΪ20��(2ǧ���֮һ��)��ԼΪ����ϸ���������������ǣ���Ϊһ������Ŀ���ת���ɴ�����������������ϸ��������Լ4000�������������д�Լ16����������Сϸ������������һ����ϸ����һ������Ƥ����ô����ϸ���ͻ��һ��18�ִ�
�����ưͿ���
�������Ȼ���ﲢ����һ�����Ŀ�������װ���˴���������װ����һϵ��������η�ĸ߿Ƽ�ϵͳ����Ŀ��ֻ��һ��:������Ѹ����Ч��������ˡ����dz�֮Ϊ��ɱ����������
�������������ġ�������ϸ������ѪҺ���ζ�ʱ�������������Լ������Ĵ̵ֶ�(1��)���ȡ�(��Լÿ��10��)����֯�л��������ߡ�������ϸ�������������롰���������ߡ�(�簩ϸ��������)�����ˣ�Ѹ�ٽ���ʶ��Ϊ���ˣ�����������װ�����ɻ���Darth
Vader����������ץס�����ɡ���Ϣ��ɱ���������������������ø�Զ��������ͼ���ܣ�����ͻ�����ᣬ����һ��ϸ���ġ����������һ���Ĵ��֣�����ס������ϸ�������β����Ƶ�ǯ�ӽ���ץס����
��һ������Ϊ�������õĹ����У�����ϸ�������ܺ��ߣ����ɲ�ʹ����Ϣ��Ȼ���������ϸ�����ڵ�һ��С����������(��Ϊ������ø��)��������ø����ڳ�һ�־��и�ʴ�Ե����ɻ���ø�Ļ�����Щ���ɻ���ø��Ѹ�ٵؽ������ɵ����ʷֽ��������ɲ���(�����ᡢ���ᡢ֬�����)������ϸ�������Щ��Ƭ����ϸ���䡰��������Ϊ�����Ͱ�ϸ���IJ�������ϸ���Ļ�����ɲ��֣������������Щ�������Ѹ�ٵض����ǽ��л������ã��Ӷ������ȫ�µĽ���ϸ����
�ҷ���������ӵġ����������ij���ÿ��������������Ϸ�����ʮ�ڴΣ�����ȫ���˾��档
����ϸ������Ҫ����������:
�
������˼�ǡ����ȡ����ǿ����Ƕ̶��ֵģ����磬����Ϊ������Ѫ���ڱڡ����ߡ��������ġ�Ȼ������������ץ�ӷ�������������������α�㣬�������Խϳ��ľ���(Ҳ��60��)������һ�£�һ����������(Volkswagen)��С�ĺ꣬�ܹ���������ϸ�����ֱ�(ÿ���ֱ۵�ֱ����Լ�൱��һ�������ܵ�ֱ��)��չ��50Ӣ����ߡ�һ������������Ǿ��ܰ��Լ�֯��һ�������Ѳ��ҵĵ�����ס��������Ŀ����һ����ϸ������ô�������һ��Ħ�г���ô�������һ��ϸ�������Ĵ�С���ͺ���Ь��ࡣ
����ϸ�����ղ�ϸ��
�������ú�������ø���γ�
һ��α���ס��������ɹ��̾Ϳ�ʼ�ˡ��������ϸ�������α���ද�����Ĥֻ�Ǽ��ں���һ�������ܺ��߾ͱ���ȫ��������ν��������ø���С�(��Phago������˼�ǡ��ԡ�����lyso������˼�ǡ�����������some������˼�ǡ�ϸ���������塱��)Ȼ��Χ������ø���Ĥ��������������������(ֻ��Ϊ��ȷ����������)����ͨ��һϵ�еĸ�ʴ��ø�����������������������ø������ݴ�����ܡ�
��Ҫ�ۿ�һ�ι��ڰ�ϸ��(������ϸ��)������ϸ���ľ�����Ƶ����������:
http://www.youtube.com/watch?v=MgVPLNu_S-w&NR=1
(����������ϸ�������Ǿ���ϸ��;����ϸ���Ĵ�С��Լ������16����)
Opsonins:������ˮ�������ǿ��������������������
Ϊ�˰�������ץס�������ϸ�������ܰ�ϸ�������źţ�ָʾ������DZ�ڵ���������Ϳһ�㱡����ճ�Ե��ס�Ȼ�����ϸ���ֱ��������ϸ���Ӵ�ʱ�����֡�������ˮ��Ϳ����Ӳ��ʹ����֮ͽ�����ѡ�
ͨ������£�һ����ᷢ��һȺ(����20������)������α������Χ���˵�ϸ��������������һ���������Ķ��������һ����������������ϸ���ͻ��������С��������ֽ�ϵIJ�Ӭһ�������˵�ϸ���ȿ�������Ҳ�������棬����û�а취���ѡ�Ȼ����������Χ�����ɣ������������ø�����ʽ����ؽ�����壬�ڱ��������ɻ���ø����������ɲ���֮ǰ�����ھ����жȹ����������ʱ�̡�
����ճ�Ե��ױ���Ϊ�������ǿ�ӡ���opsonins����ճ�ϵĹ��̽�����opsonization����
��Ȥ����,��һ����ץס�������ַ�ʽ,����Ҫ��������ʿ��֪����������,����һ��������ʿ��żȻ����һȺ�о�,���������ݵ����ʡ����������źŸ��߸����ľ���ϸ��,ʹ���������,ר�Ŵ���ר��ץ�����ĵ��ˡ��˶లȫ��(�����ϳ�Ϊ���ϵ���������ϸ���ϲ�������ı����)
��
��������ը�������ĵ�����������
����ϲ���ľ���ϸ�������ǡ���������Oxidative Blast��(Ҳ����������֪�ġ���������Respiratory
Blast��)����Ϊ������˵ĸ��ӣ����������ս��������һ�������Ǵ�˹��ά����������ߡ�λ���ͻ���Ĥ��һ��ø(��ΪNADPH����ø)�������һ�����Ե����ɵ��ӣ��������ǹ������ӵ�һ����
���ǵ���Щ������ܵľɵ��ӻ���?NADPHǹ���������ġ��ڵ��ӹܵĺ��棬����ǹ������Ļ�ϵ������ӡ�����������������ʱ����Ļ�ͻ����������γ�һ�����档ͬ����NADPHҲ���������������������ڲ��š���ã��ŵϡ�(Howdy
Doody)�������ڰ�����ϸ��������ը����Ƭ��
�������еĵ���һ��һ���س��֣������������IJ��롰���ɡ����������Ǿ����ܿ�ؾ�ȡ��һ�����ӣ��γ��ȶ��ĵ��Ӷ�(��������˵��������)�����������������ĵ��Ӿ�ȡ������Ӧ��ʹ��ϸ�����¿���ڵķ�����������������˺��С�ס����ڣ����ܺ�������һ��ı�Ĥ����ѿ��ˣ�����Ķ������������ˡ�û����������Ĥ����ϸ�������ܳ�ʱ�䡣
���������������Ƿ������ǽ��ǶԻ������˷ѡ��������ġ������㡱�ǡ����ˡ�����ϸ�������̲��������ײ�����ϸ���Ľ�����Ӵ�����һ�����롰���ˡ�ֱ�ӽӴ�ʱ�����Ż��������
���ǵ������ﵽ�����������ӡ�(������Ҫ�����������������ǣ���Щ��ȼ�ϡ������Dz���������������Դ����Щ���������ƶ������е�ϸ����ѧ��Ӧ��ʹ������Ϊ���ܡ�)�����DZ��ͷ�ʱ�������������еĴ�������ӻ�ײ����Щ�����ڵ��������е�һ��������Щ�����������ǻ�Ѹ�ٵػ������ĵ��ӣ��Ӷ��γ��ȶ��ĵ��Ӷԡ�����������ʧȥ��һ�����ӣ����ת���ǿ��ʴ�Ե����ɻ�����Ϊ���������(O2-)������������Ҫ���ӣ�Ϊ�˵õ����ӣ�����ݻ�һ�С����κζ����������������ǵĺ������ļ���ץס�IJ�����ϸ����ϸ����ͻȻ�������߷����Լ�����Ĥ����һ�������ܿ�ͻ����ġ�
���ɵ��Ӻͳ�������Ҳ������������Ӧ���γ������������ɻ�������֮һ���ǻ�����(OH-)�����ǹ������⣬�����Ǹ���ɫƿ����Ķ���һ������������Ч��������33������һ���ֲ�������ϸ����������dz��ʺϼ�ը���������ϸ����
ͨ�����������ڵ���ԭ�ӣ���������������к��Ĵ�����(HClO)�����������κ�ƽ��ĵ���Ĥ�ϴ�һ������������������һ���ж������ɻ������������Զ����ǵĵ�����ɾ�ľֲ��˺���
/ý��/ͼƬ/ hypochlorous-acid.gif
����һ�£���(������������˵)��Ϊʲô�����Լ���ϸ��û�б��Ѿ����ڻ��ƻ���?���������������������?��ΰ������⡣������һ���������ϣ���ֹ���ɵ��Ӻ����ɻ��ƻ������Լ���ϸ����������ΪSOD(���������绯ø)����һ��ר���ڳ���������������ɻ��ƻ����ǵ�ϸ��֮ǰ�к����ǵ�ø(һ�ִ��͵����ʷ���)��Ϊ�����̶ȵı�����SOD�������ھ���ϸ�����(��Ĥ)NADPH�������߲��������Աߡ�
��
��Ͱ��(NADPH����)���electron-generating��������ǹ��Ŀ���������ϸ�������һ��С����SOD���Ӱ�Χ�ͱ���,�γ�һ�֡����塱��������ǹ�;���ϸ����ϸ����ֻҪ���Dz�ͣ�����Ƥ(�������ͣ�������������ھͻ�����)�����ǾͰ�ȫ��;�����������˺����ǡ�����һ���dz�������:һ����Ц������������Ϊ�û��ṩ���õİ�ȫ�����������㣩��
��
gcmaf����ĺ��롰����������
����ǰ��������������ұ�����˵һ��:ֻ��GcMAF����ĺ���ܲ����㹻��Ч������������������Բ�����ϸ����Nagalase�ú����˯��״̬�����������ͻ��˻���һ�������˺��κ��˵�С����ǹ��������������˵ȱ�������������������������۵����⡣���ǵ���Щ������Ϊ��������Ƭ��?������һ����ͷ����ʱ���ͣ�٣�Ȼ����������һ����ͷ?����ǹ��֮����һ�κܳ��ļ�������������������ص�����������һ��ʧЧ��mac:�ڳ�ǩ��ʱ���ٶȺ�����Ͷ������Ҳ���ࡣ�ѹ�ӡ�ڰ��˻������͵��⡣������ĺ�ÿ�뷢���൱���������ӵ���ԭ���ӵ���������Զ����ͣ�������¼��ء�һЩ����ӳ�ĵ�Ӱ����̫����ӵ��Ӹ��������������������������������������ġ�����ֻ��gcmaf����ľ���ϸ�������ṩ�����ֻ�����
������ø��������(�Ͳ����)
���һ�������ϸ����ij�ַ�ʽ�������������������ú�������������ô������������֮���д�����ϸ��һ�������ɡ��ڻ���Ƕ�뵽����ϸ����ϸ�����У����˾ͱ������ھ���ϸ��(��Ϊ������ø��)�ڲ���һ��Բ�������У���������ø������������̱����䵽��������С�Ư�������鲻�ᷢ����������ø���ڲ��������ϸ����������δ������������ø��ġ������ҡ��϶��Ὣ������(�����ɡ�����˼�ǡ��Ե���)����Lyso������˼�ǡ���ɢ����"
Some "��" sack "��" bag "����˼��
һ���ֽ������ɣ�������ø�廬��������ϸ��Ĥ�Ӵ�����֮�ںϣ�Ȼ���������ķֽ����(���ᡢ֬���ᡢ�������)�ͷŵ�ϸ����Һ�С�Ȼ�����DZ�������ϸ�����գ���ѭ�����µ����岿λ�����ǵ�������̬��ʶ����Ӧ�ûᷢ��������̵�Ч����ֵ������ġ�û��ʲô���˷ѡ����µ��ж��Ļ��˱�ɱ�����������ת��Ϊ�����ĺ���:���ǡ�
���ӵ�ͨ��ϵͳ
̸̸ͨ��ϵͳ��!����ϸ����������ϸ�����ܰ�ϸ��������ͣ�غ���������һ����������������ң�ÿ���˶���ͬʱ��̸��Ȼ��������˵���ǡ���ʹ���ӡ���һ���ͷţ����������ɵ�����������ɵģ�����ϸ��ʵ���Ͽ��Ա�����˵��!!û�б�Ҫ��Թ������!�����֣����Ҷ�����������˵��İ������������ͬʱ˵���������ķ�ʽ��ÿ�α���˵��ʱ��ͣ��������˵��Ҫ��ö�(������������ͨ����������)��
������̫��Ļ�������Ų��ϵķ����ͷ���ϸ���������Ծ����ܿ���ٶ�ץס��ɱ�����˵�ϸ����������żȻ�Ĺ۲��߿��ܻ�õ����ҵ�ӡ�������ܴ���ش��ˡ�����û�аѵ�Ŭ��������ҵĽ�����һ����һ�ж���֯�÷dz��ã�Э���÷dz��á�
����ϸ������������ϸ��֮��Ļ�ѧ��Ӧ�����Ѹ�ٺ���Ч��������һ�����ӵľ���ͨ��ϵͳ����������һȺ�������绰�ĺ��ӡ����ͷŴ�������ʹ����(ϸ�����ӡ������ء�����ϩ������С����)����ÿ��ϸ��ÿ���ͷ���ǧ�����ӡ�ÿ������Я��һ���ض��������������硰��������������ߡ�������ҪһЩ�Ǹ��������ߡ�ɱ�����п������������Ķ�������������������Ҫ��֢��Ӧ�������ߡ����Dz���Ҫ����ô���ˡ������������۵��˵ij���������ԡ����Ǹ��߱˴����Ŭ�����������DZ��Ŀ���Թ�����ϸ��ʶ���ɱ��������̸�۵��˵IJ���֮�����������۵�ǰ�ĵ���ս�ԣ��Լ������õ�սʤ����
ָ�����ҿ�¡:�ռ�����
������Բ������Ҫ��һ���ǣ����������������˺꣬��ô���Ǿͻ����˿���:���ǻ�Ѹ�ٷ�ֳ�������Ƿ����Լ�����һ����ϸ���������ܶȸߵ�����ʱ�����Dz���Ҫ�ټ�����IJ���;����ֻ�ǿ�¡�Լ����ں̵ܶ�ʱ���ھ�������������ĺ���Զ�ת���������г��ĸ�����������������ǣ�����˷�����ֻ�����ڼ���ĺ��С�
GcMAF����
û��GcMAF����ͻ�˥�ˡ�����GcMAF���ڵ�����£����ǵĻˮƽ��ָ����������һ���������ͻ�Ѹ�ٷ�ֳ���������ҵĹ���������һ���У��ҽ�����Ϊʲô����
��
Chapter 6: Your Awesome Macrophage Killing Machine
http://gcmaf.timsmithmd.com/book/chapter/15/
��
GcMAF��Ϊһ�ְ�֢����
�������ư�֢��?

GcMAF��ʲô?
GcMAF��һ��ά����d��ϵ��ס�������ѧ�س�ΪGc���������ľ���ϸ���������ӡ�����һ��֧������ϵͳ�ĵ����ʣ���Ȼ�����������С�GcMAF�������ϸ����������ֿ���Ⱦ�ͼ�����ϸ����
GcMAF�Ͱ�֢
GcMAF��һ����Ȼ���������ڵ�ά���ص��ס����������֯����ϸ������������Ը�Ⱦ����֢�����߷�Ӧ�������������ɱ����ϸ����DZ����
����ϵͳ�������DZ�����������ϸ����Ⱦ��Ȼ���������֢�������γɣ���Щ����ϸ�����书�ܾͻᱻ��ϡ�
��ϸ���������ͷ�һ�ֽ���nagalase�ĵ����ʡ��������ͷ�ʱ��������ֹ����ϵͳϸ������������Ȼ��GcMAF���ױ���ֹת����һ����ǿ���߷�Ӧ����ʽ������������ϵͳ����������������������ֿ���Ⱦ�Ͱ�ϸ����
GcMAF��Ϊһ��ʵ����֢����
����GcMAF������ϵͳ�е����ã���һ��������Ϊ���ֵ����ⲿ��չ��ʽ���������ư�֢��DZ������������Ϊ��ͨ��������ע���ⲿ��GcMAF���ף�����ϵͳ���Ը��õع������Կ���ϸ����
�������Ʒ�����û�б�������ҽѧ��;�������Ǹ߶�ʵ���Եġ����һ���ٴ����������о�һ������ȻGc�������İ�֢�����Ʒ���Ȼ����û���о���������������ǵ�һ��ʹ�üȶ����о�ָ���������������Ʒ�����
ijЩ�������������������Ʒ������о��ܵ������ɡ���һ�������У�����GcMAF�Ͱ�֢���о������ء�����һ�������У�������Ϣ���о�С��Ҳ���۵����ʲ��������ˣ����������ͻ��
GcMAF�Ʒ��ĸ�����
����2002�귢���ڡ����Ұ�֢�о����ڿ����ϵ�һƪ����GcMAF�����£����ܴ���GcMAF��С�������û�С����Ի�����֢�������á�
��ǰ����ʲô?
��Ϊһ�ֿ�����Ч�İ�֢�����ֶΣ�GcMAF�Ʒ������о��С�Ȼ������Ҫע����ǣ�GcMAF�������û�б����������ư�֢�������������⡣
�������������ͳ�İ�֢����ѡ���ѡ��GcMAF�Ʒ��������о��������ԣ������й���GcMAF���ư�֢��������ֵ�û��ɵġ���ijЩ����£��о���ԱΪ��������ҩ��Ĺ�˾����������������£���Щ�о��������ֱ����ء�
����Ҫ���н�һ�����о����ڴ�֮ǰ��GcMAF�ڰ�֢�����е��κ��������ö��Dz�ȷ���ġ�
GcMAF and Cancer: Get the Latest Information
https://www.healthline.com/health/gcmaf-cancer-treament#outlook
��
��
����ϸ���;ƾ���صĸ�����֢
Cynthia Ju��ʿ��Pranoti Mandrekar��ʿ��
Cynthia Ju��ʿ�ǿ��������ѧ�����ҽѧУ��˹����˹ҩѧԺ�Ľ��ڡ�
Pranoti Mandrekar��ʿ������������ѧҽѧԺҽѧϵ�Ľ��ڡ�
������о���������ϸ���ھƾ��յ��ĸ�����֢�ķ�չ�����Źؼ����á�Ϊ��ȷ����ƾ��Ըβ�(ALD)����Щϸ�����²����ܣ����ٴ������н��й㷺���о��Խ�һ������ALD�¾���ϸ���ı��Ͷ������Ƿdz���Ҫ�ġ�����Ϊֹ���о��Ѿ�ȷ���˾���ϸ���ļ����ص㣬��������ϸ���ļ����ͱ��Ͷ����ԡ������ķ�����Ҫ�����ھ���ϸ��/����ϸ���Ľ�����פ�����Լ�����ϸ������������������Ӫ��������չ�е����á��о������ʾ����ϸ�����ܳ�Ϊ�ƾ��Ը����˵����ưе㡣δ�����о����������˽�ƾ����Ӱ������������֯�ľ���ϸ������ת�������������Ƿ��������ϸ����ALD�еĹ��ܣ��Լ�����ϸ���İ����Ƿ��ܼ���ƾ��Ը����ˣ���Щ����Ϊ�ƾ��Ը����ߵ������ṩ��ϣ���IJ��ԡ�
�ؼ���:�ƾ�����;�ƾ��Ըβ�;�ƾ��Ը�����;�ƾ��Ը���;�ƾ��η���;��;������;�������߷�Ӧ;��Ӧ�����߷�Ӧ;����ϸ��;����ϸ������ת��;����ϸ��
�ƾ��Ըβ�(ALD)��һ��Ӱ��ȫ���������˵ĸ��Ӽ��������տɵ��¸�Ӳ���ΰ�(����Ӳ��)����ϸ����)�����˾ƾ������л����ȩ�Ը�ϸ����ֱ��ϸ�����Ժ�����Ӧ���鵼�����⣬����ƾ����ἤ�������������Ӧ�����߷�Ӧ����Щ��Ӧ�漰���ָ�ϸ�����ͣ�����פ������ϸ������Ȼɱ��ϸ������Ȼɱ��Tϸ�����ܰ�ϸ����������ϸ�����ر��ǣ������еľ���ϸ����Ҳ��Ϊ���ո�ϸ����������������еIJ�ԭ�壬����ϸ�������������߸������Ϊ�ڶ��ػ�֬����(LPS)��ϸ�����������ϸ���dz���Ҫ(Jenne��Kubes
2013;��ķɭ��ŵ��(2010)������������������λ���ཹ��������ת����/����С��ģ�ͺ���һ��������о�����ʹ���Ǹ��õ��˽����ϸ������ѭ��ǰ��ϸ��(������ϸ��)����Դ�������Ժͱ��ͺ��ܵĿ����ԡ�����ϸ��)��
����ѭ������ϸ���ļ���;���ϸ���ľۼ���ALD������Ҫ�IJ�������������Ȼ�����ξ���ϸ����ALD���������е�������δ��ȫ���������Ľ����۵���ϸ��/����ϸ������ѧ�����һЩ�·��֣�����Ŀǰ���ڸξ���ϸ��������������Ӫ�������е��о���չ����ȷ���д���������⣬�Կ����������������Ӫ�������ľ���ϸ���������ơ�
����ϸ���;���ϸ�����ͺ��ܵ�������
����ϸ����Ҫ��Ϊ������:�����Ծ���ϸ������֯פ���Ծ���ϸ��������ϸ���ڻ���Բ�ԭ����������˵ij�ʼ��������Ӧ��������Ҫ���á�����ľ���ϸ�������ڳ�Ϊ����ϸ����ǰ��ϸ��������ϸ��������ѭ��������֢��Ӧ����ʱ������ϸ�������յ���֯�С����֮�£���֯פ���ľ���ϸ����ʼ�վ�����һ����֯�ڣ��䵱�ڱ��͵�һ�����ߣ���������֯�е��κθ�Ⱦ�����ˡ�
��
����ϸ�������ڸ�����֯�У�����ԡ�Ƥ�����Ρ��κ�Ƣ����֯פ���ľ���ϸ��������פ����֯���ֳ����ֱ��ͺ��ܣ�����������Щϸ����Դ������(Davies et
al.
2013)�������֬����λ�о���ʾ����̥�ѻ��Һ�/��̥������ǰ��ϸ����������֯פ������ϸ������Դ��������Щ�ڸ����еľ���ϸ����Ƥ����������ϵͳ(Gomez
Perdiguero��Geissmann 2013)����֯פ���ľ���ϸ��������Ϊһ�������Ե�����ϸ��Ⱥ����ά���ض���֯����̬���ܷdz���Ҫ(Davies et
al.
2013)����֯����ϸ�������Ҹ��»��Ǵӹ����в��ϲ�������һ������������⡣Ȼ��������֤�ݱ�����������Դ��ѭ������ϸ����������֯��֢���ڱ���ļ�����˲�λ����������ֻ�Ϊ����ϸ�������䵥��ϸ���ͷǾ��䵥��ϸ���������˵�����(���Ⱦ����)��������֯��˳����ļ�����⣬�ݱ�����פ���ľ���ϸ����ļ���ĵ���ϸ�����ܹ���ijЩϸ������(���ϸ������(IL)-4)���յ��½������Ҹ���(Jenkins�ȣ�2011��)��
�����еĿ��ո�ϸ������֯�ھ���ϸ�����������ģ��Ǹ�����֢��Ӧ����Ҫ��Դ��Kupfferϸ�����������߹��ܲ����ڻ���Ը������˵ķ�Ӧ����ؼ����ã������ڸ���Կ�ԭ�����鷴Ӧ����ؼ����á����ո�ϸ��λ�ڸ���Ҫ��Ϊ������Ⱥ(Klein
et al. 2007):
�������еľ���ϸ��Ѹ�ٱ���Ѫǰ��ϸ��ȡ��������֢��Ӧ������Ҫ����;��
��������֢����ͷ��䡢���ٵĿ��ո�ϸ����
С��ģ�ͳ��������о�����ϸ�����ܵĸ������档Ȼ�����뵥��ϸ��ǰ��һ������ʹ��С����Ϊ�������ٴ�ǰģ��ʱ�����뿼��С����������ϸ�������IJ���(Mestas��Hughes
2004)�����磬С����������ϸ���ڱ���Ϊtoll������(TLRs)�ı�����ӵı����Ͽ���������ͬ��TLRs�������ϸ���ļ������������ķ�Ӧ�Լ�����һ��������
����ϸ������
����ϸ����һ�ֶ��ص��������ı�����ͺ��ܣ���ȡ������֯��������ʾ����ϸ�����ӡ��������ӡ���ԭ����ط���ģʽ����(PAMPs)��������ط���ģʽ����(DAMPs)�Ĵ��ڡ�������̱���Ϊ���������������־���ϸ�����͵ij��֣��ֱ���ΪM1��M2����ϸ����M1����ϸ����Ҫ���д������á����磬������M1����ϸ�������鵼��ϸ����ϸ���Ͳ����ij�ʼ����;���⣬���Ƕ���֯���˵ķ�ӦҲ����Ҫ��M1����ϸ����������ѹ�����ʺ�ϸ������,��il
- 1�������������Ӧ�(TNF��)���ø����ء�����,����,һ������,�ͻ�����(ROS),���ܼ�����Ӧ�����߷�Ӧ(Jouanguy et al .
1999;Shaughnessy��Swanson 2007)��һ����Ⱦ�����˱����ƣ�����ϸ��ת��Ϊ���ס���֯�����ͣ��Կ��ƹ��ȵ���֯������֢��Ӧ(Benoit
et al.
2008;Noel���ˣ�2004)����Щϸ��ͨ������Ϊ���������ϸ��(M2),�����ٽ���֢����֯���ľ���(�����Ͷ�����ĺ�2012)(��ͼ1)�����ǿ�������M1����ϸ���Ĵ��ڸ�ˮƽ�ļ�����ǵ�����(��Fizz1,
Mrc1 Ym1, __arg1)(���2003;Mantovani�ȣ�2002)��

ͼ1����ϸ�������Լ��������֯���˵�ʾ��ͼ������֢��Ӧ����ʼ�Σ�����ϸ����ļ�������˻��Ⱦ�IJ�λ����M1���͡�������������֢��ѹ�����ʺ�ϸ������,�������������Ӧ�(TNF��)������(IL) 1��-12��,�����ئ�(IFN��),ø����һ������(��ӱ���),�ͻ�����(ROS)����Щ����ϸ�����д��Ϳ������ã������»��ʽ������֯�ƻ����ھ���ε�����,��ЩM1����ϸ��ת����һ��ƽ���ױ��Ͳ�ͬ��ϸ�����Ӻ���������,il - 10��ת���������Ӧ�(TGF-��)�����ʽ�������ø(MMPs),������ø1 (__arg1),��֯��������ø���Ƽ�(TIMPs)��Ѫ����Ƥ��������(VEGF)����ЩM2����ϸ�����п������ã��ٽ�Ѫ���γ�(Ѫ������)�����ʺϳɺ���֯���ܡ�
��
����ϸ���Ĺ��������Է�ӳ�����ڸ��ּ����еIJ�ͬ(��ʱ�෴)������(Sica��Mantovani
2012)�����磬��ȻM1ϸ���ڼ��Ը�Ⱦ�ڼ�����ϸ���Ͳ���������Ҫ����M1��Ӧʧ�����ܵ��¸�����֯���ˡ���ˣ�M1����ϸ���Ĵ����ܵ������������Լ���(��ؽ��Ͷ��Ӳ��)�ʹ�л�Լ���(���ȵ��صֿ������Ͷ�������Ӳ��)�����Ƶأ���ȻM2����ϸ��������֯�������ߵ�����أ��������M2��Ӧ�ɵ������Լ���������Ӧ��Ƥ�ס���������֯��ά�������⣬����ʱ������ƣ�������֯�����ı仯�����µ�ϸ�����ı�ļ���Ҳ���ܰ����ž���ϸ�����͵ĸı�(������ϸ�����͵ĸı�)������ϸ��������)�����磬�ڰ�֢���ڣ�������صľ���ϸ�������ھ��伤���M1ϸ�������Դٽ����������߷�Ӧ��Ȼ�������������Ľ�չ����Щ��������صľ���ϸ��ת��Ϊ���ر��ͣ����ƿ��������߲��ٽ���������(Allavena
et al.
2008)����һ�������ǣ��Ƿ��ָ����е�֬����֯����ϸ����Ҫ����Ϊ�������ϱ��ͣ����ٲ�������ϸ�����ӡ�Ȼ�����ڷ��ֻ����У�֬����֯����ϸ��ת��Ϊ������m1�����ͣ���������ϸ�����ӵIJ��������ȵ��صֿ�(Zeyda��Stulnig
2007)��
��Ȼ������ϸ����ΪM1��M2ϸ���ܷ��㣬����Ҫע����ǣ����ֻ��ֹ��ڼ�M1��M2����ϸ��������ȫ�ױ��͵��������ˣ��������������ж��о��в�ͬ���ͺ��ܵ���Ⱥ�����磬M2ϸ�����Է�Ϊ������������:�˿������ͺ����ߵ����;���ϸ��(Edwards
et al.
2006)���˿����ϵľ���ϸ���ڸ������͵İ�ϸ���ͷŵ�ϸ������IL-4��IL-13���������γɡ���M1����ϸ����ȣ���Щϸ�������Ĵ���ϸ�����ӡ�ROS��һ������ˮƽ�͵ö࣬���ٽ���֯�������˿����ϵķ���ˮƽ�ߵö�(���¶�����塢ϸ������ʳɷֺ͵��ڻ����ؽ�������)���෴�����ߵ��ھ���ϸ����������Ӧ�����߷�Ӧ�ĺ��ڻ�Ӧ���յ�����Ƥ�ʼ����ϵ��ķ�Ӧ�С���Щ����ϸ�����ص��Dz�����ˮƽ��IL-10���յ����ߵ��ھ���ϸ�����������Ӱ������߸����ǰ�����ء�����ϸ�������ա��鰷��֬���ء����˿����Ͼ���ϸ����ͬ�����ھ���ϸ�����յ�ϸ��������ع���
��
����ϸ�����͵Ŀ������ܶ���ϸ���ڷ��ӻ��ƿ��ƣ������źŵ��ס�ת¼���Ӻͱ����Ŵ��¼������磬����ϸ��ͨ��TLRs����������ļ���յ���һ���漰STAT1��2���ӵ��źŻ��ƣ�ʹ����M1���ͷ���(Qin�ȣ�2012��)���෴��ͨ��IL-4/IL-13��stat6�鵼�Ļ��Ƶ�ѡ���Լ������M2����(Daley�ȣ�2010;Ī��ŵ�ȣ�2003;Stolfi et al. 2011)������m2��������ͨ��IL-10/STAT3��IL-3/STAT5�źŻ����յ���(Sica��Mantovani 2012)������ϸ����������һ����Ҫ����������ʹSTAT6���ữ��JNKø(Shirakawa et al. 2011)��ȱ��JNK�����MLK3�ķ���С��ȱ��M1����ϸ����������ʾJNK��M1���͵ļ�����������(Gadang�ȣ�2013��)������ijЩ����ת¼��IRF����Ҳ�Ǿ���ϸ����������Ҫ�������ӡ����磬IRF5�Ļ��Դٽ���IL-12�����ת¼����M1������أ���IRF5���������յ���IL-10��������M2����(Krausgruber et al. 2011)�����Ƶأ�����һ����ΪNotch/IRF8�ĵ��ڵ�������ᵼ��M1����(Xu et al. 2012)��������M-CSF/IRF4�ᵼ��M2��������һ�౻��ΪSOCSs�ĵ�����Ҳ���ž���ϸ����������Ҫ�������ã��������Ե�ϸ�����Ӵ̼���SOCS���Ͳ��������ϸ���Ƿ����M1��M2���͡����,il - 4�Ĵ���������SOCS1 / STAT1�շ�M1����(Whyte et al . 2011��),�������ئ���TLR�����յ�SOCS3 / STAT3�͵���M2����ϸ������(��ŵ�¡�et al . 2014��)����������λ��ϸ����ϸ����,����ӳ�ΪPPAR��PPAR��,Kruppleϲ��4,��ԭ������Ҳ���¾���ϸ�����������������ڽ�����/ STAT-SOCSͨ·(��et al . 2014��)������ڲ��ı�DNA���е������Ӱ��DNA�ṹ�ͻ������ĵ��ع��̡��ٽ�M2���͵��յ�������m1��������(Banerjee et al. 2013;Satoh���ˣ�2010)����Щ�����Ŵ��������Ӱ����鵰��ȥ��ø��Jumonji D3��microRNA let-7c��
��
��ALD�У�����ϸ��ӡ�Ǻ�M1��M2���͵ļ����ܵ�������ϸ�����ӽ��ʵ�Ӱ�졣��ALDʱ������ϸ�����Ӻ�Ӧ���������ͨ·����ϸ�о����������������˽���ھ���ϸ���ļ�����
����ϸ�����˻�
����ϸ����ALDӪ�������е�����
����ϸ���ƺ�������������Ӫ�������������������á���ʵ�ϣ�������о������������ALD�в�ͬ���͵ľ���ϸ��������ӵ������(Lee et al.
2014)����ˣ��Ծƾ���֬���Ը����ߵĸ�����������������֯��ѧ�����������˱��������ϸ�����ӵľ���ϸ������Щ�����ϸ������ͨ����M1ϸ����أ�ͬʱҲ��M2ϸ����ء�����������������������ϸ�����ܿ�����ƾ��Ը�����ά�����ߵļ���״̬�����ٴ�����ԣ���������:
����ϸ���������������������ڶ��б���(����֬����)������(��(Karakucuk et al. 1989)�����ܾ���ϸ�������뼲�����س̶�֮��û����ȷ������ԡ�
����ϸ���������Ӳ�����Ƹ��ˮƽ,������MCP-1 MIP-1��,MIP-1��,���ӻ��ߵĸ���ALD (et al . 1998��)��
�ڻ�����������У���֢�����ھƾ���ظ�Ӳ�����߾���ϸ���еı�����ڱ����ײ�����ظ�Ӳ�����߾���ϸ���еı���(Tapia-Abellan et al.
2012)��
����ϸ��������أ����µ��ʺͰ�ϸ��������ؿ�ԭ3��������������Ӫ����������������(Luna-Casado�ȣ�1997��)��
��ALD����ѭ������ϸ��������Է�����TNF��TNF�����塣��LPS�̼�ʱ,�����ͷų�����ˮƽ��TNF��(Gobejishvili et al .
2006;�ŵȣ�2001)���߶�TNF��ˮƽ����ѪҺ��,������,��ƶ���йؽ���ڼ��Ծƾ��Ը�����(��et al .
1990��)����ijЩ�����,�����Ŀ���ϸ������il - 10ˮƽ��δ�����ƹ�������TNF��(Le Moine et al . 1995��)��
�ƾ��Ը���/���Ӳ�����ߵ�����ϸ������(��IL-6��IL-8��IL-18)��ѭ������ϸ���ξ���ϸ����������������ˮƽ����(Afford et al.
1998;Fisher���ˣ�1999)����Щ���ӵ�ϸ������ˮƽ���ٴ�������(Khoruts et al. 1991;��������Ϳƶ�1989)��
�ƾ���ظ�Ӳ�����߸���������ȫ������������ʾ�˶��صĻ������ģʽ���ڸ�Ӳ�������ں����ڸ�����ͬ����Ӳ�����ڽ����ڸ߱���Ļ�����������ϸ�������ֳ��Ǩ����صĻ���(Lederer
et al. 2006)��ǿ������ϸ����ALD��չ�е����á�
��Ҫ������ٴ��о���������ͬ��ALD���ߵľ���ϸ����ѭ������ϸ�������˽ⵥ��/����ϸ���Լ�����չ��ȷ�й��ܹ��ס�
��
Kupfferϸ����ALD�е�����
Kupfferϸ����פ���ڸ���ľ���ϸ����ͨ��CD14/TLR4���帴���D���Ӧ�Գ������ƹ�����LPS�ij���λ���ӣ�����ܵ��¾ƾ��Ը����ˡ������о���ʾ,���Ժ������Ҵ�������CD14��TLR4�ļ���ľ���ϸ���ڸ���,����upregulation
TNF��CD14�����Ӳ���,MCP-1, ROS(�\��ʧ��et al .
2001��)�����⣬ͨ������;������������ϸ������Ԥ���ƾ��յ��ĸ�����֢(Koop�ȣ�1997;Petrasek��(2012)��֤ʵ���յ���������Ҫϸ����
�о���Ա�Ѿ���������������μ�����ո�ϸ���ġ�ROS�IJ��������Ǿƾ��Ը�����Kupfferϸ����LPS�����Ļ���֮һ(Thakur et al.
2006)���ڳ�ʱ��Ӵ��ƾ��Ĺ����У�Kupfferϸ������ROS��������ͨ��һ�ֲ������ƾ���л��ø���յ��������ġ�(Kono et al.
2000)�������������ڿ��ո�ϸ����еĹؼ�����Ҳ���о��еõ���֤ʵ��NADPH����ø)�����ֹ�һ��Ԥ����ROS����alcohol-fed�����Լ������źŷ��ӵ����ữERK1/2�����������Ĵ���ϸ������TNF������ϸ��(��Ұet
al . 2000;Thakur���ˣ�2006)��
�ھƾ��鵼�Ŀ��ո�ϸ����У���һ����Ҫ�ijɷ���CD14/TLR4���帴���lps�յ���Kupfferϸ�����������帴����ļ���ᴥ�������źż�ø(�������˹�����IKK),���յ��´���ϸ�����ӵ��յ�TNF��,il
- 6,
MCP-1���������ģ����,����ϸ��alcohol-fed���������ϻ��˺�չ�����ϻ������ӷ�Ӧ����,���¸���ˮƽ��TNF��(��2003)��MCP-1
(Mandrekar et al . 2011��)������TLRs����ǿ����Ҳ�ɴٽ�ros�鵼�ľƾ��Ը�����ո�ϸ������(Gustot et al.
2006)��Hritz����ͬ��(2008)��Inokuchi����ͬ��(2011)֤ʵ��TLR4������������Ӫ���������ߵ�Kupfferϸ��������Դ������ϸ���ϱ������Ҫ�ԡ�Ȼ����Ŀǰ������������е�kupffer
-cell������TLR4�Ƿ���Ψһ���¾ƾ��鵼�ķ������Ƶ�TLR�����������Ҫʹ��ȱ������ϸ��������TLR4��С����н�һ�����о���Ȼ������ĿǰΪֹ���о�����������ƾ��յ���ROS��kupfferϸ�����ڶ��ص�����������(�ڶ��ؿɵ��´���Ӧ��ǿ)��ALD��kupfferϸ�������Ҫԭ��
����Kupfferϸ���Ļ�ͼ��ٴ���ϸ�����ӡ����ر�������Kupfferϸ����������ϸ�����ӡ����Ѿ���Ϊ����ALD��һ���ص㡣���磬����ͨ��ʹ��֬����(һ�ֿ���֬������)�����ƾ���¶��kupfferϸ��ԭ������������תkupfferϸ������(Thakur
et al. 2006)������֬������״��ֹLPS-stimulated TNF������ͨ���������ϸ��il - 10 / STAT3 /
hemoxygenase-1ͨ·���յ�M2����ϸ��(2011��Mandal et al .
2010��)����������M2����ϸ���ƺ�����ٻ����ĸ�������أ���Ϊ��Ŀǰ��ȸ����˺�֬�����Ե��������У�M2����ϸ��ռ������λ�������ظ����˵Ļ��������ΪM1����ϸ��(Wan
et al.
2014)����һ�ֿ��ܵ�ALD���Ʒ�����ͨ������IL-10ˮƽ�����;ƾ���¶��Kupfferϸ���������ԡ��ƾ��յ���IL-10�����ѱ�֤�������ھ���ϸ����������IL-10ȱ��С����о����־ƾ��鵼�Ĵ�����ϸ�����ӵIJ�������(Hill�ȣ�2002��)��������о�Ҳ������il
-10ͨ������ƾ��Ը����е�TLR3��������(Byun et al.
2013)��ѡ���Եذ���Kupfferϸ���е�TLR�ź�ͨ·���ܻ������������Ӫ���������ߴ���ϸ�����ӵIJ���ƽ��Ĺ����ṩ���õļ��⡣
3 .����TLR3��TLR4�⣬�����黯���ھƾ��Ը����߸������з���TLR2��TLR8��������(Lee et al. 2014)��
ALD�иν������ϸ��
��֯פ���ľ���ϸ����������еĿ��ո�ϸ���������ܵ�����ԭ�壬���ܰ���������ά��ϸ��(���ϸ��)��ȷ����֯��ƽ�⡣Ȼ������û�и�Ⱦ������£��ɸ�Ⱦ����֢�����Ӧ��������(�����⣬����ϸ������������֯���ֻ�Ϊ����ϸ�������������ԭ�壬�������ϸ����ϸ����Ƭ���ָ���֯�ڻ����ȶ�����ʵ�ϣ������༲��ģ����(�縹Ĥ��֢)���ѱ���������֯����ϸ��ʵ������Դ�����ֽ���ĵ���ϸ��(Ghosn
et al. 2010)��
���Ժ����Ը����˵��о�Ҳ֤ʵ�˵���ϸ���ĸ��ھۼ������磬�����Ȼ�̼(CCl4)��С����м��Դ����ᵼ�¸������ˣ����¾���ϸ���Ĵ���ӿ�룬ʹ�������ϸ����������10��(Karlmark�ȣ�2009��)�����һ�������ccl4�յ��ĸ���ά��С����о�����������ϸ���Ľ�������ά���Ľ�չ�������з�������Ҫ����(Ramachandran
et al. 2012)��ͬ�����ڶ������������յ��ĸ�����С��ģ���У��ָ�����ļ�ľ���ϸ���������֯������Ҫ����(Holt et al. 2008)��
���Ծƾ��Ըβ�Ҳ������ͨ������ϸ������鵼�ʹ����ģ���Ϊ���Ծƾ���ҩ�ɵ���С������н����Ծ���ϸ���Ļ���(Wang et al.
2014)������ľ���ϸ������������Ⱥ- ly -
6chi��Ly-6Clowϸ�����������Ե��Ŵ�������Ly-6Clowϸ�����ֳ�������֯�������ͣ������ˮƽ�Ĵ���ϸ�����Ӻ�ˮƽ�Ŀ��ܲ�����֯���Ŀ�����(Arnold
et al.
2007;Nahrendorf�ȣ�2007)���෴��Ly-6Chiϸ������Ϊ������֯���˱���;Ȼ���������ɵ����ĸ�ϸ���������ƺ�ת��ΪLy-6Clow����(Wang
et al.
2014)����������Ⱥ�Ľ������ϸ�����棬����������֢��֯����ģ���б��ֳ����Եġ���ʱ�෴�Ĺ��ܡ�����������ģ���У�����Ly-6Chi����ϸ������ļ�����˵����࣬��������ֻ�Ϊ�������Ե�Ly-6Clowϸ��(Lin
et al.
2009)����ccl4�յ��ĸ���ά����������У�������Ly-6Chiϸ����Ly-6Clow�������ϸ���Խ����֢����ά�����ָ���֯��̬������Ҫ����(Ramachandran
et al.
2012)�����ļ������н���-����ϸ����Ⱥ�����õ��о�Ҳ������Ly-6Chi��Ly-6Clowϸ��������ļ����֯������ϸ��Ly-6Chi���������ڽ�����֯�����е���ˮ������ɹ��ܡ����ļ������ĺ��ڣ�ļ����Ly-6Clowϸ�����м�������֢���ԣ���ͨ���ٽ�Ѫ���γɲ�����֯�������ļ�ϸ���ļ���(��Ѫ������)��(Nahrendorf�ȣ�2007)��
�������У�����������(���Ǿ���CD14+CD16+����ϸ����Ly-6Clow�������ϸ�����Ӧ���ڶ�����֢�Լ����з������������ʪ�ؽ��ס���������Ӳ������������Ӧ��ʪ������ͷ��ݵ��׳���֢���Ǿ����CD14+CD16+����ϸ��Ҳ�����Ըβ����ߵ�ѭ�����������ţ���ʾ���Dz����˸�����֢����ά���Ľ�չ(Ziegler-Heitbrock
2007)��
ALD�еľ���ϸ������
�����д��ڵķǾ��ʺͽ����Ծ���ϸ�����ж�����ALD��صĹ���(��ͼ2):
���ǿ�����Ϊ��ԭ�ʵ�ϸ�����ڱ�����ʾ�������ӣ��Ӷ�������Ӧ�����߷�Ӧ��
���ǿ��ܱ��ֳ�������ȩ-��ȩ���ι��ĸε��Ĵ��ڡ�������ȩ-��ȩ�Ӻ���)(Willis�ȣ�2002)���������λ�ı�������ʵĹ��ܡ���ALD�����У���Щ�Ӻ���Ҳ��������������Ĵ����йء�
����ͨ�����������ĺͽ��ʣ�����������ɱ�����;Ȼ������Щ���ܿ���������������Ӫ�������ڼ��ܵ���
ͨ������tlr�鵼���źţ����ǿ��ܵ���Tϸ���ϵ�������������PD-1��TIM-3�ı������ӣ��Ӷ������ƾ��Ը����ߵĿ�������(Markwick et al.
2015)��
ijЩ��Ⱥ(��Ly6Chi�������ϸ��)�������ִ����ʣ����������������Ե�������ϸ�����Ӻ��������ӣ��Ӷ�������֯���ˡ�
ALD�еľ���ϸ������
�����д��ڵķǾ��ʺͽ����Ծ���ϸ�����ж�����ALD��صĹ���(��ͼ2):
���ǿ�����Ϊ��ԭ�ʵ�ϸ�����ڱ�����ʾ�������ӣ��Ӷ�������Ӧ�����߷�Ӧ��
���ǿ��ܱ��ֳ�������ȩ-��ȩ���ι��ĸε��Ĵ��ڡ�������ȩ-��ȩ�Ӻ���)(Willis�ȣ�2002)���������λ�ı�������ʵĹ��ܡ���ALD�����У���Щ�Ӻ���Ҳ��������������Ĵ����йء�
����ͨ�����������ĺͽ��ʣ�����������ɱ�����;Ȼ������Щ���ܿ���������������Ӫ�������ڼ��ܵ���
ͨ������tlr�鵼���źţ����ǿ��ܵ���Tϸ���ϵ�������������PD-1��TIM-3�ı������ӣ��Ӷ������ƾ��Ը����ߵĿ�������(Markwick et al.
2015)��
ijЩ��Ⱥ(��Ly6Chi�������ϸ��)�������ִ����ʣ����������������Ե�������ϸ�����Ӻ��������ӣ��Ӷ�������֯���ˡ�
��
����ϸ�����Ӻ����������⣬����ϸ�����������ʻ���������ϵͳ��֬���غ����ص�֬���ӡ�����C3��C5ϵͳ��С��ALD�����ڱ�����ϸ��������ٽ������ķ����ͷ�չ(Roychowdhury
et al.
2009)���෴��֬�������ƺ���С������������Ӫ�������ľ���ϸ�������и��������á���ˣ��б�ҪѰ���µľ���ϸ����������������������Ӫ�������ļ������Ӷ�Ӱ���䷢չ��
���Ÿ������˵Ľ�չ������ϸ��Ҳ��Ҫͨ�����������������ϸ����ϸ����Ƭ�����dzɹ������֢�ʹٽ���֯���Ĺؼ����衣����ϸ������������,��ʼ��������ϸ������,��il
-
10��TGF-��(��ɭ�Ͳ�����2013;Korns����2011��;�Լ����д������õ��������Ӻ���֯���ܽ��ʡ����,������֯��ѧ�����һ���о��۲���ǿ��TGF-�±���ʽ�ľ���ϸ���ƾ��Ը����ߵĸ�������(Lee
et al .
2014��)��Ȼ�����������˳�������ʱ��������֢����֯�����̿ɵ�����֯��ά������֯����������(����������һ�����е�һ�����أ���Ϊ�������Ҵ�ι����۲쵽������֯ȱ��(Arteel�ȣ�1996��)��ȱ�����µ����ʵ��ȶ��ͼ����Щ�����ʱ���Ϊȱ���յ�����(HIFs)�������ڶ���;��������ϸ������ֳ�ʹ�л������ϸ�������ۼ���ȱ����������֯��(Murdoch
et al. 2005)��С��ģ�͵ĸ����ˡ����Ը������յ��ľ���ϸ������HIF1��,�ٽ���ά��ͨ����������pro-fibrogenic����(����et al .
2012;Mehal 2012)��
����Ӧ���鵼�ľ���ϸ����Լ���������Ӱ�����ϸ��������ϸ��������ALD��֢����Ҫ����
ALD�����ơ�������ϸ����һ�����ܵ�Ŀ����?
���ۼ�����չ��ʲô�Σ����һֱ�������������˻�֢����Ч�����Ʒ�����Ȼ������������ȱ�������������ԣ����¸�������һ�ַ�������ʹ��п(Kang��Zhou
2005)��ά���غ�s
-���յ��������ָ��ƾ��Ը�Ӳ�����ߵ�Ӫ��״̬�����������������ޡ�ˮ�ɼ��غ���˼������Ʒ�Ҳ����������ALD��δ���ٴ�����(Frazier�ȣ�2011��)���������ư������ϸ������,������Ƥ�ʼ�������,pentoxyfylline,��anti-TNF������,Ҳ������ALD����30���ꡣȻ��������Ϊֹ���ɹ������ġ��ڼ��Ծƾ��Ը�������ʹ����Ƥ�ʼ��ص��ٴ�������ʾЧ�������ԣ���������Ϊ��Ѫ֢����������Ѫ�������Ӷ���ֹ(Maddrey
et al.
1978)���������о��������Ƶ�Ч�����ض�anti-TNF������,�ٴ������ijɹ�����ˣ�Ϊ�ƾ��Ը���Ӳ�������ƶ���Ч���Ե�������Ȼû�еõ����㡣
Ϊ��������һ�����о���ԱҲ������������Ծ���ϸ���IJ��ԡ���Щ����ͨ��ʹ��ϸ���������Ƽ���ϸ���ڽ���������ϸ�����ӵIJ�������һЩ��ϣ���Ľ��:
����������Ƶ���IL-1���ź��ھ���ϸ��ʹ��il - 1��������(��anakinra)ȡ���˼������Ƶ��µ�С��������֢��Ӧ(Petrasek et al .
2012��)��
������״֬����ͨ��Ѫ��������ø-1�յ�Kupfferϸ������IL-10���о�������С��ALD (Mandal et al.
2010)��ͨ�����ڸ�����һ����̼�Ŀ����������յ��ξ���ϸ���е�����ø��ALDС��ģ����Ҳ�����������(Bakhautdin et al. 2014)��
Χ��Kupfferϸ����С��ƾ��Ը����н������ϸ��������MCP-1��MIF���о�������Щ������������Ч�İб�(Barnes et al.
2013;Mandrekar���ˣ�2011)��
���Ӧ���յ������ݿ˵���90���ض����Ƽ��IJ��ԡ���Ŀǰ�ڰ�֢�ٴ�������������һ�ַ�������ͨ������С��������ϸ����֢��Ӧ����������ALD (Ambade et
al. 2014)��
��Щ�о���֧ͬ���˾���ϸ�����������ھƾ��Ը������е��ٴ����ۡ���Ծ���ϸ�����ܺͱ��͵������Ƽ���С����ҩ��Ϳ������������ϵ��ٴ��о�����Ϊ�ƾ��Ը���Ӳ���������ṩ�־õ���Ч��
���ۺ۵�
�������������Լ���һ��������ϸ���ѳ�ΪALD�Ĺؼ������ߣ������ܳ�Ϊ����Ŀ�ꡣȻ����������Щϸ���������Ժͱ��Ͷ����ԣ��ľ����еĸξ���ϸ��������һ����Ч�ķ���;�����������ض���Ⱥ���д�ȷ����
���ڸξ���ϸ����ALD��չ�е����ã���������������Ҫ��������磬ALDʱ���ڽ���ĵ���ϸ����ηֻ�?��ʲô��֯�����źźͷ����ź�ͨ·�����˾ƾ��Ը��о���ϸ�����ٱ��?��һ����Ҫ�������ǣ�����֯�ڻ����ָ��ȶ�����ο��ƽ������ϸ���������������ϸ����������ǻ�Ǩ��?���⣬�����˽���ƺ͵��ڸξ���ϸ�����ͺ��ܵķ��Ӻ�;�����ƶ�����ALD�IJ���������Ҫ�����磬��ALD���ڣ�Ly-6Clow�������ϸ������֯�����˿������еĹ��ܿ�������Ԥ�����Ը�����֢����֢��֯���˵�Ly-6Chi�������ϸ��������֯����Ly-6Clowϸ����ת��������Ϊ��������ALD�İе㣬��ƾ���֬���Ը��ף������������С���о������(Singal
et al.
2013)�����ƾ���ϸ���鵼����֢��Ӧ�Ѿ����������������µ�����ѡ��;���磬��͡��ҩ��������ͪ��ҩ���n-3֬����ȿ���Ԥ������ϸ���鵼����֢��������������ѡ����(Ji
et al. 2009;Methe et al. 2005;Ramirez�ȣ�2008;Yeop Han et al.
2010)����Щ���Ʒ����ڼ���ƾ���֬���Ը������˺���֢�����Ч��Ҳֵ�����ۡ�
��
Phenotypic and Functional Heterogeneity of Monocytes and Macrophages
Macrophages, which play an important role in the initial innate immune response
to infection with pathogens or other insults fall into two main
categories��infiltrating macrophages and tissue-resident macrophages.
Infiltrating macrophages are derived from precursor cells called monocytes that
circulate throughout the body and are recruited into the tissues when an
inflammatory reaction occurs. Tissue-resident macrophages, in contrast, always
remain localized within one tissue, serving as sentries and first line of
defense against any infection or injury in that tissue.
Alcohol Research: Current Reviews | Macrophages and Alcohol-Related Liver
Inflammation
https://www.arcr.niaaa.nih.gov/arcr372/article09.htm
��
����ϸ���鵼��֢�������������������е�����
.jpg)
ͼ1��ʾ��
M����Ӧ��Ƥ���˿ڡ�trM��s����������Դ:�ѻ�������̥��̥�����ࡣ��Ѫ���÷�������Щλ��,��ϸ��,������֯F4/80hi
M��s������������ļ�ĵ���ϸ��Ⱥ�����ڹ��裬Ȼ���ͷŵ�ѪҺ�С���֯���˺�,����������ϸ������������ļ��վ�ڵ�������Χ��֯�ͷֻ���MoM��s��Ly6Chi
(M1) MoM��s�ͷŴ�����,��Ly6Clo (M2) MoM��s���ڿ����ʡ�GMP, granulocyte-M����;��Ѫ��ϸ��;MDP,
M��-dendriticϸ���档
ͼ2��
Stage-dependent M��s���˿����ϵĹ��ܡ�ROS����������

ͼ3��
�˿����ϵı����Ŵ��仯���鵰������(�������ͼ���)��DNA������RNA�����dz����ı����Ŵ�ѧ�ı䣬������Ӱ���˿������ڼ���֢�Ļ������仯��ء�
Macrophage-Mediated Inflammation in Normal and Diabetic Wound Healing | The
Journal of Immunology
https://www.jimmunol.org/content/199/1/17/tab-figures-data
��
Anti-oxidant inhibition of hyaluronan fragment-induced inflammatory gene expression.
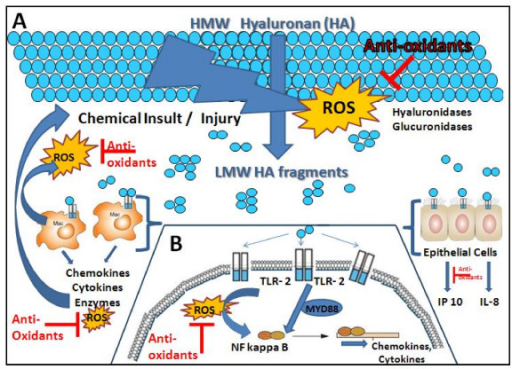
F6: ROS�ڲ�������������ѭ�����е���������:a)��֯���˲���ROS���鵼ϸ�������HA�����顣B)��ɢHA��ROS��ǿͨ��TLR-2������������ϵͳ,�����ROS�IJ���,��һ������NF-��B������ϸ�����Ӻ��������ӵı���,��һ���ٽ���֢�����ѭ��ʹϸ�������HA�ij�����Ƭ����ROS�����ɣ��Ӷ����½�һ�������ˡ���֢�����յ���ά�������������п��ܸ������ֶ���ѭ����
��
The central role of ROS in a pathophysiologic "vic | Open-i
https://openi.nlm.nih.gov/detailedresult?img=PMC2627834_1476-9255-5-20-6&req=4
��
����������֢����֯�����е�����
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3929010
��������(ROS)�IJ�����������֢������չ�Ĺؼ���ROS�ɲ�������������Ӧ��ϸ������������κ�������ϸ��(PMNs)����ͨ���ؼ���ϸ���źŵ���(���Ұ�������ø)�������ٽ���Ƥ�����ϰ���
Reactive Oxygen Species in Inflammation and Tissue Injury
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3929010
Mar 01, 2014 �� Production of reactive oxygen species (ROS) is central to the
progression of many inflammatory diseases. The ROS are produced by cells that
are involved in the host-defense response, such as polymorphonuclear neutrophils
(PMNs) and promote endothelial dysfunction by oxidation of crucial cellular
signaling proteins such as tyrosine phosphatases.
��
����������֢����֯�����е�����
������(ROS)������֢�Լ�����չ������Ҫ���õĹؼ��źŷ��ӡ���֢��λ�Ķ��κ�������ϸ��(PMNs)��ǿROS���ɣ�������Ƥ�����ϰ�����֯���ˡ�
����������֢����֯�����е�����
www.ncbi.nlm.nih.gov pmc /����/ PMC3929010 /
Reactive oxygen species (ROS) are key signaling molecules that play an
important role in the progression of inflammatory disorders. An enhanced ROS
generation by polymorphonuclear neutrophils (PMNs) at the site of inflammation
causes endothelial dysfunction and tissue injury.
Reactive Oxygen Species in Inflammation and Tissue Injury
www.ncbi.nlm.nih.gov/pmc/articles/PMC3929010/
��
��
���������������������������cell.com
https://www.cell.com/cancer cell/fulltext/s1535 - 6108 (14) 00470 - x
ֵ��ע����ǣ��������ƹ����ĺ����������Ŀ�����;���ɵ��������Ⱙϸ����Эͬ������������������ֿ���������������չ����Ҫ�ԣ�������Ϊ���Ƹ�Ԥ��DZ�ڰе㡣
Glutathione and Thioredoxin Antioxidant ... - cell.com
https://www.cell.com/cancer-cell/fulltext/S1535-6108(14)00470-X
Remarkably, combined inhibition of GSH and thioredoxin antioxidant pathways
leads to a synergistic cancer cell death in vitro and in vivo, demonstrating the
importance of these two antioxidants to tumor progression and as potential
targets for therapeutic intervention.
�����������������͡���Ӧ�е�����
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC543461
��HDACi��������ϸ����ת��ϸ�����������յ�ת��ϸ����������ϸ���ĵ������������Ȳⶨ��SAHA�������˷γ���άϸ��WI-38����sv40ת��������VA13��Ӱ�졣����(��2.5��M)����ϸ��������WI38
VA13ϸ��(ͼ1 A��C)��
Role of thioredoxin in the response of normal and ...
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC543461
Jan 18, 2005 �� HDACi Arrests Cell Growth of Normal and Transformed Cells and
Induces Apoptosis of Transformed but Not Normal Cells. We first determined the
effect of SAHA on normal human lung fibroblasts, WI-38 cells, and its
SV40-transformed derivative, VA13. SAHA (�� 2.5 ��M) inhibited cell growth of WI38
and VA13 cells (Fig. 1 A and C).
��
����������Դ
ϸ����ROS��һ����Ҫ��Դ��NADPH����ø��NADPH����ø��O2��NADPH���ɳ��������Щø�����������ϸ���б����ֵģ�������ϸ���У�����ͨ���ھֲ�������ˮƽ������Ӧ��[9]��ɱ�������ɵIJ�ԭ�塣����һ�����������Ѿ��۲쵽NADPH����ø�����Ա�����ڻ����������֯�У���Щ��֯�Է����߹���Ҳ����Ҫ[10,11]��ר�Ų�����������ø�Ĵ���֤ʵ�˻�������ϸ������������õ�ģ�ͣ������Ǽس䵱�ж��ĸ���Ʒ�����⣬�°�������Դ̼�NADPH����ø������ROS���������ѱ�֤����ϸ����ֳ[12]������ġ�NADPH����ø��ϸ���ڶ�λ������ϸ������������Ĥ��ϸ���ˡ������������������Ȥ���ǣ����������Ҳ��֤����NADPH����ø��������ROS��[13]����ȻNADPH����ø��ϸ����ROS��������Դ����������ܣ���ƪ��������������������Դ��ROS�Ļ��ƺͺ����
ϸ��ROS����������������塣�ݹ��ƣ��������ܺ�������1%���ڲ�����������[14,15]���������а˸���֪���ܹ��������������λ��[16,17]����Щλ���ϸ����ROS����Թ����в������Ȼ�������Ը�����I��II��III��ROS����֤����ϸ���ź�ת��[16]��Ӱ�졣��Ȥ���ǣ�������I������II��������������ͷ�ROS��������III������������������Ĥ[18]�����ͷ�ROS���������Ͻ������������ͷŵ���Ĥ�ռ佫�����ӽ����ʰС�����һ������һ�µ��ǣ�����iii������ROS�ѱ���ȷ֤������������ѧ��������Ҫ����������Ӧ��ϸ���ֻ�����Ӧ������[19]��mROS��������Դ�Ƿ������mROS�ź�ת������ͬ�����в������Mitochondrial
reactive oxygen species and cancer | Cancer & Metabolism | Full Text
https://cancerandmetabolism.biomedcentral.com/articles/10.1186/2049-3002-2-17
��
Cited by: 6
Publish Year: 2013
Author: Federico S. Rodr��guez, Katterine A. Salazar, Nery A. Jara, Mar��a A
Garc��a-Robles, Fernando P��rez, Lu...
Vitamin C in Stem Cell Reprogramming and Cancer
https://www.cell.com/trends/cell-biology/fulltext/S0962-8924(18)30064-3
Sep 01, 2018 �� However, elevated basal levels of ROS, deficiency in catalase
activity or increased uptake of vitamin C by tumor cells could render them
selectively vulnerable to the pro-oxidant effect of high-dose vitamin C, and is
an ongoing area of research.
Cited by: 17
Publish Year: 2018
Author: Luisa Cimmino, Benjamin G. Neel, Iannis Aifant
��
ά����C�ڸ�ϸ���ر�̺Ͱ�֢�е�����
ά����C������������Ҫ����������ά����Cת�����ϸ����Ѫ��ˮƽ����ά����Cת���帺����ʳ���պ�ȫ�����ա�
ά����C��һ�ֿ�����������2 + /��-KGDDs��������ʽ��
ά����Cͨ������JmjC���鵰��ȥ����ø�ͷ۷������ǻ�ø�Ļ��ԣ���ǿ����άϸ����iPSCs�ı����ر�̣���ά��ESC�Ķ����ԡ�
ά����Cȱ����ͨ������TET�ǻ�ø���ԣ��ٽ���Ѫ��ϸ���쳣���Ҹ��£����ٰ�Ѫ����չ��
ҩ��������ά����C�ɻָ�����״ϸ���ķ۷�����ܣ���Ѫ����������֢�ı������нϺõĸ������á�
ά����C������Ļ�����ʳ��������Ϊһ�ֿ�������,ά����C��һ����������ʽ��2 +,��-ketoglutarate-dependent��˫��ø(Fe 2 +
/��-KGDDs)����������ͬ��ø,������ԭprolyl�ǻ�ø�ͱ����Ŵ��ļ�ܻ������鵰��DNA������ά����C���Ե�����̥��ϸ��(ESC)����,����������άϸ�����յ����ܸ�ϸ��(����),���谭���쳣������Ѫ��ϸ��(hsc)ͨ����������ߵĻҪôJumonji
C (JmjC)
domain-containing�鵰��demethylases��һǧ��һʮһ��λ(����)DNA�ǻ�ø�����ڱ����Ŵ�ʧ������֪�Ķ����������������أ�ά����C������Ϊ�����Ŵ������������µ����á�
�ؼ���
����C
�����Ŵ�ѧ
��ϸ��
���ڵĵ�����
����
����ά����C������������Ҫ����Ѫ��ˮƽ����������ά����Cת�˵����ϸ���ƣ���Щת�˵�������ʳ���պ�ȫ�����ա�
ά����C��һ�ֿ�����������2 + /��-KGDDs��������ʽ��
ά����Cͨ������JmjC���鵰��ȥ����ø�ͷ۷������ǻ�ø�Ļ��ԣ���ǿ����άϸ����iPSCs�ı����ر�̣���ά��ESC�Ķ����ԡ�
ά����Cȱ����ͨ������TET�ǻ�ø���ԣ��ٽ���Ѫ��ϸ���쳣���Ҹ��£����ٰ�Ѫ����չ��
ҩ��������ά����C�ɻָ�����״ϸ���ķ۷�����ܣ���Ѫ����������֢�ı������нϺõĸ������á�
ά����C������Ļ�����ʳ��������Ϊһ�ֿ�������,ά����C��һ����������ʽ��2 +,��-ketoglutarate-dependent��˫��ø(Fe 2 +
/��-KGDDs)����������ͬ��ø,������ԭprolyl�ǻ�ø�ͱ����Ŵ��ļ�ܻ������鵰��DNA������ά����C���Ե�����̥��ϸ��(ESC)����,����������άϸ�����յ����ܸ�ϸ��(����),���谭���쳣������Ѫ��ϸ��(hsc)ͨ����������ߵĻҪôJumonji
C (JmjC)
domain-containing�鵰��demethylases��һǧ��һʮһ��λ(����)DNA�ǻ�ø�����ڱ����Ŵ�ʧ������֪�Ķ����������������أ�ά����C������Ϊ�����Ŵ������������µ����á�
�ؼ���
����C
�����Ŵ�ѧ
��ϸ��
���ڵĵ�����
����
��֢
Vitamin C in Stem Cell Reprogramming and Cancer: Trends in Cell Biology
https://www.cell.com/trends/cell-biology/fulltext/S0962-8924(18)30064-3
��
2-DeoxyGlucose (2-DG) - Blog: Cancer Treatments - from ...
https://www.cancertreatmentsresearch.com/2-deoxyglucose-2-dg
Oct 24, 2015 �� Summary: This is a glucose analog that is avidly taken up by
cancer cells. DeoxyGlucose (2DG) differs from normal glucose only by removal of
an oxygen atom from the hydroxyl group at the 2 position. 2DG is one of the most
relevant glycolysis inhibitor. Since it is a glycolysis inhibitor, it is
relevant to all cancers visible on PETContinue reading 2-DeoxyGlucose (2-DG)
��
2-����������(2-DG) -��־:��֢����-���ԡ�
https://www.cancertreatmentsresearch.com/2-deoxyglucose-2-dg
���ܽ�:����һ�ֱ���ϸ��̰�����յ�������ģ�������������(2DG)�����������ǵ����������2λ�ǻ��ϵ���ԭ�ӱ�ȥ����2DG������ص��ǽͽ����Ƽ�֮һ����������һ���ǽͽ����Ƽ�������������PETContinue�Ͽɼ��İ�֢��أ���2-����������(2-DG)
��
Biology of glucose metabolization in cancer cells ...
https://www.sciencedirect.com/science/article/pii/S2452336417300420
After the discovery of based on the altered cancer cell metabolism in 1930,
loads of studies have shed light on several aspects of cancer metabolism with a
common goal to find new ways for effectively eliminating tumor cells by
targeting their energy metabolism.
����ϸ�������Ǵ�л����ѧ��
https://www.sciencedirect.com/science/article/pii/S2452336417300420
1930�꣬���ڸı�İ�ϸ����л�ķ��֣��������о���ʾ�˰�֢��л�ļ������棬��ͬ��Ŀ�����ҵ��µķ�����ͨ����������ϸ����������л����Ч����������ϸ����
����:13
������:2017
����:Adewale Fadaka, Basiru Ajiboye, Oluwafemi Ojo, Olusola Adewale, Israel
Olayide, Rosemary Emuowhocher��
Cited by: 13
Publish Year: 2017
Author: Adewale Fadaka, Basiru Ajiboye, Oluwafemi Ojo, Olusola Adewale, Israel
Olayide, Rosemary Emuowhocher...
TargetingTumor MetabolismWith 2-Deoxyglucose in Patients ...
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4142700
Sep 15, 2010 �� DISCUSSION. This is the first study to define the recommended
phase II dose, pharmacokinetics, and molecular marker data for 2DG, a
prototypical inhibitor of glycolysis, as a single agent. The ability to use 2DG
in the clinic lays an important foundation in the effort to target metabolism in
cancer.
2-���������ǰ���������л
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4142700
2010��9��15�������״ν��ǽͽ�ĵ������Ƽ�2DG��Ϊ��һҩ����ȷ���Ƽ���II�ڼ�����ҩ������ѧ�ͷ��ӱ�����ݵ��о�����2DG�����ٴ�Ϊ�����İ����л�о��춨����Ҫ�Ļ�����
����:151
������:2010
����:Mark Stein, linhongxia, Chandrika Jeyamohan, Dmitri Dvorzhinski, Murugesan
Gounder, Kevin Bray, Kev��
Cited by: 151
Publish Year: 2010
Author: Mark Stein, Hongxia Lin, Chandrika Jeyamohan, Dmitri Dvorzhinski,
Murugesan Gounder, Kevin Bray, Kev...
A Phase I/II Trial of 2-Deoxyglucose (2DG) for the ...
https://clinicaltrials.gov/ct2/show/NCT00633087
Mar 11, 2008 �� A Phase I/II Trial of 2-Deoxyglucose (2DG) for the Treatment of
Advanced Cancer and Hormone Refractory Prostate Cancer (2-Deoxyglucose) Fasting
blood glucose < institutional ULN. In the Phase I portion of this study patients
may have had prior chemotherapy. In the Phase II portion of this study patients
may not have had prior chemotherapy. Women of childbearing potential must have a
��
2-����������(2DG)��I/II�����顣
https://clinicaltrials.gov/ct2/show/NCT00633087
��2-����������(2DG)�������ڰ�֢�ͼ���������ǰ���ٰ�(2-����������)��I/II�������ڱ��о��ĵ�һ�Σ����߿����Ѿ��й�����ʷ���ڱ��о��ĵڶ��Σ����߿���֮ǰû�н��ܹ����ơ�������DZ���ĸ�Ů�����С���